





1 引言
蛋白质作为生命活动的核心执行者, 通过动态的分子间相互作用形成具有时空特异性的多分子组装体, 在时序性激活级联生化反应通路中发挥决定性作用, 从根本上塑造细胞的表型特征[1]. 这些多分子复合物受到精密调控, 介导了细胞通讯[2]、信号转导[3]、物质运输[4]、表观遗传调控[5]以及代谢稳态维持[6]等关键生物学过程, 而异常的蛋白质-蛋白质相互作用(Protein-Protein Interaction, PPI)与神经退行性疾病、恶性肿瘤等重大疾病的发生发展密切相关[7-9]. 因此, 系统性解析PPI, 不仅有助于揭示生命现象的本质规律, 更能为精准医学时代的药物研发提供新视角与新思路.
作为解码信号转导网络与复杂生命系统的关键切入点, 全面绘制PPI网络图谱已成为现代分子生物学研究的重要方向. 为此, 研究人员构建了多维度的技术体系, 包括免疫共沉淀[10]、酵母双杂交[11]、荧光共振能量转移[12-13]、荧光/发光蛋白互补[14]等. 然而, 这些方法普遍依赖于融合表达标签蛋白质, 对于目标蛋白质的功能可能存在一定影响, 并且外源表达的融合蛋白质无法精确反映天然蛋白质的丰度和后修饰水平. 近年来, 基于质谱的蛋白质组学[15-16]方法为高通量检测PPI提供了新策略, 通过整合亲和纯化[17-18]、尺寸排阻色谱[19-20]或邻近标记[21]等富集技术, 研究者已实现对天然细胞裂解液中多种PPI的系统性鉴定. 但该策略仍面临双重挑战: 一, 其检测灵敏度受限于样本量需求(通常需大于106个细胞), 难以适配临床来源的稀有细胞类型(如循环肿瘤细胞); 二, 基于群体细胞的平均化检测模式, 丢失了单细胞维度的互作网络异质性信息.
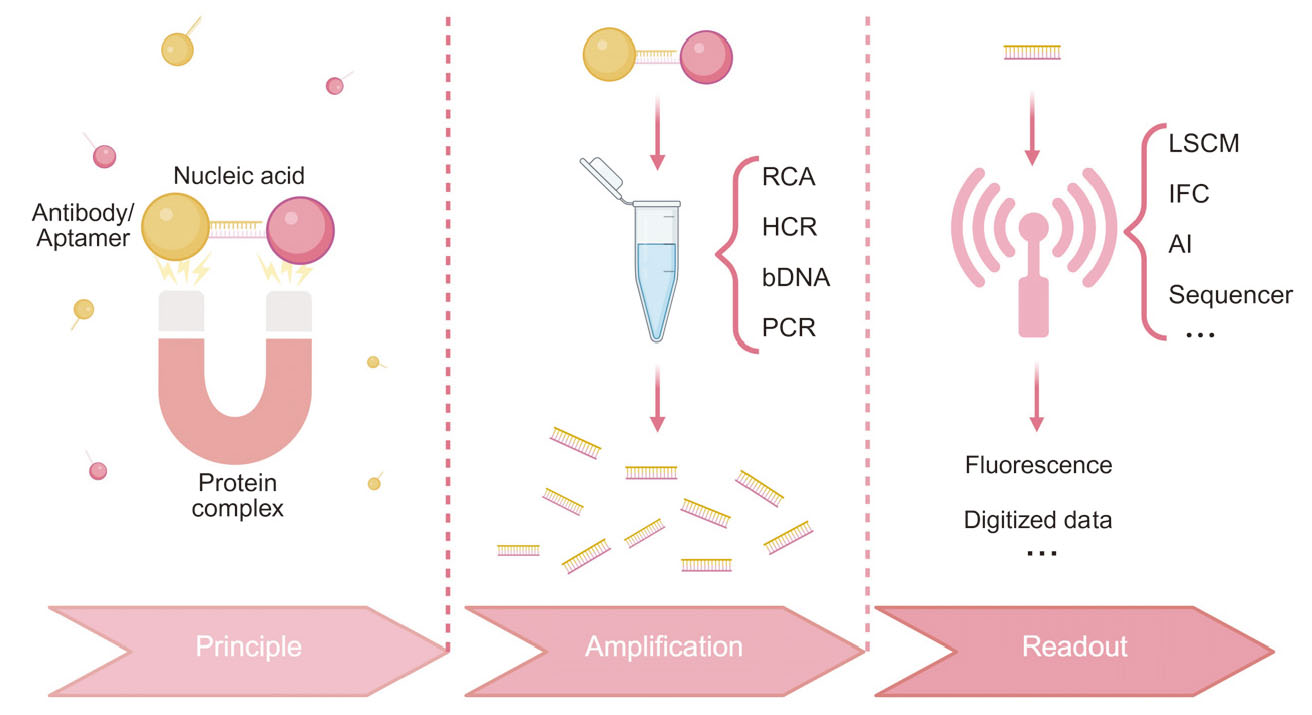
DNA纳米技术用于PPI检测的核心原理在于空间邻近依赖性[26-27]的自组装反应, 其功能性构架可分为三个模块: 传感模块、信号放大模块和信号读出模块. 传感模块通过抗体-DNA复合物等修饰有单链DNA探针的配体分子高亲和力、高特异性结合靶标蛋白质; 当靶标蛋白质之间存在相互作用时, 邻近抗体上的DNA探针序列触发空间邻近响应性自组装, 从而将PPI事件转化并记录为可扩增的核酸序列. 信号放大模块依托于酶促或非酶促的核酸扩增反应, 对序列化信号进行放大, 获得在光学或高通量检测中足够被捕获的信号, 以实现低丰度蛋白复合物的高灵敏检测. 信号读出模块则通过荧光显微成像、流式细胞术或高通量测序等手段, 并辅以人工智能算法, 对放大后的核酸序列信号进行定性或定量分析, 完成对PPI事件的数字化解析.
本综述将系统性阐释邻近依赖性核酸自组装在PPI分析中的设计逻辑. 首先聚焦于PPI诱导的局部浓度富集效应, 剖析其如何驱动相应DNA探针的空间重构与自组装; 继而评述基于滚环扩增(Rolling Circle Amplification, RCA)、杂交链式反应(Hybridization Chain Reaction, HCR)等的信号放大体系策略, 重点解析其在PPI检测中的优势及应用瓶颈; 在此基础上, 深入探讨传统光学检测平台的性能优化途径, 以及基于核酸测序平台的数字化读出方法所带来的多重检测与高通量优势. 最后, 展望该领域与人工智能、微流控芯片等前沿技术的跨学科融合, 为精准医学和新药研发带来创新范式变革.
2 邻近依赖性核酸自组装用于PPI检测的基本原理
蛋白质分子在发生特异性相互作用时, 其中心距离通常缩小至10 nm以下. 如此纳米级的空间特征, 为邻近依赖性核酸自组装技术提供了关键的局部浓度富集效应, 使得分子间相互作用能够被高灵敏度地捕捉, 完成PPI事件向可读出核酸信号的转变.
邻近依赖性核酸自组装技术的检测原理基于空间限域效应引发的局部浓度质变[27](图1A): 当两条分别偶联于单克隆抗体上的互补DNA探针游离于溶液中时, 其杂交行为受限于极低的分子碰撞频率. 在自由扩散状态下两条DNA探针的平均间距为微米量级, 难以形成稳定的双螺旋结构. 当单克隆抗体结合相互作用的两种蛋白质时, 两条DNA分子共定位于同一分子复合物的表面, 其有效活动范围急剧缩小至抗体-靶标蛋白复合物决定的纳米级空间. 这种空间限域效应使得DNA探针的局部有效浓度提升超百万倍, 极大增加了两条DNA相互碰撞并发生杂交的概率和速率, 也显著增强了双链结构的热力学稳定性(图1B). 研究者在设计此类临近依赖性DNA自组装序列时, 应确保在未结合相互作用蛋白时因有效浓度低而难以形成稳定的双链, 而在结合后可快速稳定杂交. 这需要综合考量DNA序列的长度与碱基组成, 以确保核酸自组装过程严格受控于PPI事件.
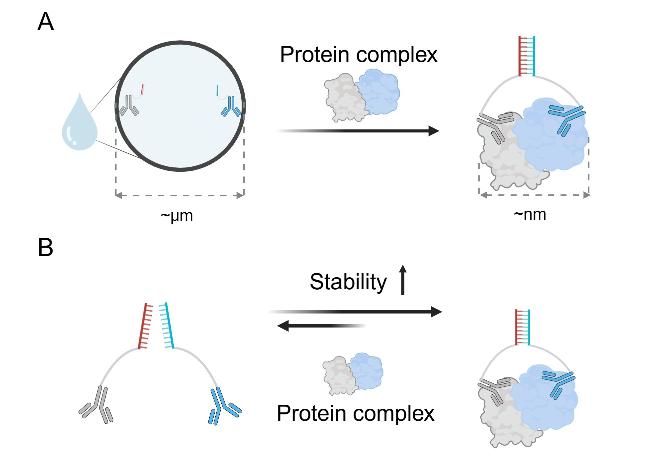
基于上述原理, 该技术可将PPI信息转化为稳定的核酸双链信号, 继而通过体外核酸扩增实现信号放大, 并结合荧光成像或高通量测序等多元检测平台, 最终达成单分子水平的PPI网络解析.
3 信号放大技术
在邻近依赖性核酸自组装技术中, 信号放大模块是实现内源性PPI高精度解析的关键. 通过构建酶促或非酶促核酸扩增反应, 该模块能突破传统检测方法的灵敏度瓶颈, 实现对低丰度PPI的超高灵敏检测. 目前, 常用的信号放大策略主要包括RCA、HCR以及聚合酶链式反应(Polymerase Chain Reaction, PCR)等. 本节将系统阐述上述信号放大技术的机理原理, 并探讨优化扩增效率与提高信噪比的发展方向, 以期为绘制PPI图谱提供指导性思路(表1).
表1 不同信号放大方法用于PPI检测的比较Table 1 Comparison of different signal amplification methods for PPI detection |
| 信号放大方法 | RCA | HCR | bDNA | PCR |
|---|---|---|---|---|
| 核心机制 | 酶促等温扩增 | 非酶促等温自组装 | 非酶促多级探针杂交 | 酶促热循环指数扩增 |
| 成本 | 中 | 低 | 中 | 高 |
| 商品化试剂盒 | 有 | 有 | 无 | 无 |
| 操作复杂度 | 中 依赖两次酶促反应, 相 对简单 | 低 无酶, 仅需1~2轮孵育 和洗涤, 反应条件简单 | 高 无酶, 但需要3~4轮孵育和 洗涤, 对操作者的要求较高 | 中 依赖至少两次酶促反应 及专用热循环仪 |
| 误差来源 | (1) DNA连接酶活性波动 (2) Phi29聚合酶活性波动 (3)高丰度时的空间信号重叠 | (1)非特异性发夹自组装 (2)反应环境中盐离子浓 度等影响组装效率 | (1)多轮孵育、洗涤操作导致 误差 (2)探针的非特异性杂交 | (1) DNA连接酶活性波动 (2) PCR的序列偏好性、 保真度 |
3.1 滚环扩增
RCA[28]作为一种酶促等温核酸扩增技术, 其核心机制依赖于环状DNA模板的结构特征. 当引物特异性结合环状模板后, 在具有链置换活性的DNA聚合酶(如Phi29 DNA聚合酶)的作用下进行持续滚动复制, 生成包含数百至数千重复单元的线性串联重复序列. 该技术凭借其高扩增效率、温和的反应条件, 尤其适用于PPI的高灵敏检测.
2006年, Söderberg团队[29]首次将RCA应用于PPI的原位检测, 提出了邻近连接原位扩增技术(Proximity- Ligation In Situ Amplification, P-LISA). 该方法利用偶联了DNA探针的特异性抗体结合靶标蛋白质. 如图2所示, 当蛋白间发生相互作用时, 其关联DNA探针因空间邻近(<40 nm)触发DNA连接反应, 形成环状扩增模板; 随后, 在Phi29 DNA聚合酶催化下进行RCA反应, 形成长串联重复DNA序列; 最终通过荧光探针杂交实现信号可视化. 研究团队运用P-LISA成功解析了内源性c-Myc/Max异源二聚体的核内动态分布, 将检测空间分辨率提升至30 nm, 显著提高了PPI检测的灵敏度与定位精确度. 然而, 传统P-LISA技术在实际应用中仍存在两大关键瓶颈问题: 其一, 微米级RCA扩增产物(直径约0.8 μm)在高靶标浓度条件下易发生信号重叠现象, 导致单分子信号难以区分; 其二, 高丰度蛋白的邻近效应(蛋白间距<40 nm)易引发非特异性连接事件, 干扰真实互作信号的判读.

针对P-LISA技术的信号重叠问题, Söderberg团队[30]在2011年创新性地设计了三浓度梯度DNA环系统(1∶10∶100)并结合多通道荧光检测, 成功扩展了P-LISA的动态检测范围, 从而在乳腺癌组织的HER2表达异质性解析中实现了单细胞分辨率的精准检测. 对于高丰度蛋白引发的邻近效应假阳性, 该团队[31]进一步在2022年开发了分子布尔逻辑门系统(Molecular Boolean Logic Operators, MolBoolean), 其核心机制在于“AND-NOT”三色荧光编码探针: 当且仅当两个探针同时结合至同一蛋白复合物时, 触发AND逻辑门信号, 表征PPI; 若仅单个探针结合游离单体蛋白(NOT逻辑门), 则输出其他荧光信号. 该技术被成功应用于TGF- β1诱导的上皮-间质转化模型, 首次实现E-cadherin/β- catenin复合体解离过程中单体动态变化的时空分辨定量, 验证了其在高背景干扰下解析动态PPI的特异性优势. 值得注意的是, 柯荣秦团队[32]于2024年提出的杂交增强邻近连接技术(Hybridization-enhanced Proximity Ligation Assay, HPLA), 通过构建双功能探针, 实现了靶标蛋白丰度与相互作用的同时检测, 为原位PPI研究提供了全新的技术手段.
目前基于P-LISA技术的PPI检测, 已成功实现了商业化, 但受限于荧光光谱重叠, 仍存在检测通量低的局限性. 为突破这一瓶颈, Coskun团队[33]于2024年开发了迭代式擦除邻近连接技术(iterative erasure sequencing- PLA, iseqPLA). 该技术通过创新性地整合DNase I/Nuclease P1核酸酶介导的探针擦除策略与多轮次荧光成像系统, 实现了单样本中22种PPI的同时检测. 研究团队应用该技术系统绘制了非小细胞肺癌细胞亚细胞分辨率下的动态PPI图谱, 首次在单细胞尺度揭示奥希替尼耐药过程中ERK-YAP信号通路的补偿性激活机制, 为克服第三代EGFR酪氨酸激酶抑制剂耐药提供了潜在的联合治疗靶点. 作为空间组学领域的前沿工具, iseqPLA独特的原位探针再生设计完整保留了PPI的空间分布信息, 为系统性解析复杂PPI网络提供了创新方法.
近年来, 基于RCA的信号放大体系通过多次迭代升级, 实现了检测通量从单一PPI到22种PPI的跨越性提升, 并在临床组织切片分析中展现出卓越的应用潜力, 其亚细胞级空间分辨能力为精准医学研究开辟了新维度. 然而, 当前技术体系仍面临两大关键瓶颈: 其一, 酶促反应体系中DNA连接酶、Phi29 DNA聚合酶以及核酸酶活性的波动可能引入系统误差; 其二, 实验周期较长且缺乏成熟的自动化平台, 限制了其在更大规模、高通量应用场景中的推广与普及.
3.2 杂交链式反应
作为典型的DNA链置换型等温扩增技术, HCR[34]的核心在于多种发卡结构DNA分子的级联自组装. 在无引发剂条件下, 发卡结构依靠茎部的互补配对维持热力学稳态; 当加入特异性引发剂后, 即可依次触发各级发夹的开环与链置换反应, 驱动DNA分子按照预先设定的程序化路径自组装, 最终生成携带荧光标记的线性双链DNA纳米结构, 从而实现高效的信号放大.
2015年, Söderberg团队[35]首次将HCR与邻近依赖性核酸自组装技术相结合, 开发出邻近依赖性杂交链式反应检测系统(proximity-dependent Hybridization Chain Reaction, proHCR). 如图3所示, 该体系利用分别偶联了发卡结构探针PH1与PH2的两种特异性抗体结合靶标蛋白复合物; 随后通过加入激活剂寡核苷酸链, 触发PH1探针的构象重排并释放桥接序列; 该桥接序列在邻近效应作用下高效激活邻近的PH2探针, 暴露隐藏的引发剂序列进而启动HCR扩增. 随后, 荧光标记的H1/H2发夹探针交替组装, 生成线性荧光双链DNA产物. 研究团队将该技术成功应用于E-cadherin/β-catenin复合体的原位互作检测, 并在甲醛固定石蜡包埋组织等复杂样本中展现出良好兼容性. 2024年, Pierce团队[36]进一步通过设计正交性邻近探针与特异性HCR序列, 成功实现α/β-tubulin、β-catenin/E-cadherin及SC35/SON三组PPI的同时成像.
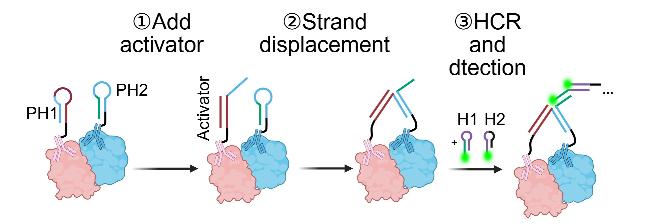
值得关注的是, 杨黄浩与李娟团队[37]在2018年开发了基于核酸适体的邻近激活HCR成像系统, 利用靶向c-Met受体的核酸适体作为亲和配体, 结合HCR信号放大技术, 在活细胞膜表面及斑马鱼模型中实现了受体二聚化的动态监测. 这种免转染、免酶的实时可视化策略, 为细胞表面PPI研究提供了创新工具.
HCR技术凭借其等温扩增特性与无需复杂温控设备的优势, 为PPI研究提供了便捷性工具. 近年来, 该技术通过多维度优化持续拓展应用边界: 将识别配体从传统抗体替换为物理尺寸小的核酸适体, 显著提升组织穿透性并实现活体动态成像; 通过热力学稳定性优化发夹探针, 在生理盐浓度下维持高杂交效率; 发展多色编码正交探针系统, 扩展同步检测通道, 支持多PPI的原位分析. 然而, DNA分子对微环境的高度敏感性以及非特异性的发夹自组装可能导致假阴性与假阳性信号, 限制了其在低丰度PPI检测中的应用. 针对这些挑战, 亟需结合人工智能辅助的核酸探针设计以及其他跨学科方法, 以进一步提升HCR在PPI研究中的精度与通量.
3.3 分支放大
支链DNA(branched DNA, bDNA)技术[38]作为一种非酶依赖型信号放大策略, 通过多级杂交反应构建稳定的三维分支骨架, 实现核酸信号的线性放大. 与PCR等指数扩增方法相比, bDNA通过构建稳定的三维分支结构逐级传递信号, 展现出显著的低背景噪声和出色的定量精确度, 其检测灵敏度可达单细胞、单分子水平. 自 1990 年代问世以来, bDNA技术已广泛应用于病毒和细菌感染的临床诊断.
为拓展bDNA技术在PPI研究中的应用, Yang团队[39]于2019年创新性地提出了分支邻近杂交技术(Branched Proximity Hybridization Assay, bPHA). 如图4所示, bPHA利用偶联了寡核苷酸探针的特异性抗体或核酸适体特异性结合靶标蛋白; 当蛋白间发生相互作用时, 寡核苷酸探针通过Z型结构的桥联核酸探针与预放大器核酸探针形成稳定的初级杂交骨架; 随后, 通过结合放大器核酸探针形成分支网络结构; 最终, 荧光标记探针结合于分支上, 实现信号输出. 该放大机制可带来高达400倍的信号增益, 结合流式细胞术或共聚焦显微成像, 能够在单细胞水平进行PPI的定量分析. 应用该技术, 研究团队成功捕捉到B细胞表面抗原受体在抗原刺激下的微簇重组现象.

bPHA技术通过线性放大机制克服了指数扩增的非线性偏差, 可在单细胞水平解析免疫细胞亚群的功能异质性, 并且模块化的探针设计使其兼容流式与显微成像等多种检测平台. 然而, bPHA依赖多步探针孵育与精密洗涤, 对实验操作的规范性与技术人员的熟练度提出了较高要求.
3.4 聚合酶链式反应
PCR作为分子生物学的基石技术, 通过热循环介导的DNA变性 、引物退火和酶促延伸三阶段反复循环, 实现对目标序列的指数级扩增. 该过程可将单拷贝DNA分子扩增至106~109拷贝, 为痕量核酸检测提供了关键技术支撑. 定量PCR (Quantitative PCR, qPCR)通过引入荧光报告系统(如SYBR Green双链嵌入核酸染料或序列特异性TaqMan探针), 能够实时监测扩增动力学并实现精准定量[40], 已成为基因表达谱分析、病原微生物检测等领域的标准化方法.
2024年, Shao等[41]在《Nature Biotechnology》上报道了基于PCR与DNA邻近连接技术的新型PPI分析平台TETRIS (Tandem Elongation of Templated DNA Repeats for Analysis of Interacting Proteins). 如图5所示, 当靶蛋白在三维空间内发生相互作用时, 偶联在特异性抗体上的含6 nt粘性末端的双链DNA在T4 DNA连接酶的催化下发生邻近连接反应, 生成记录有相互作用信息的线性DNA, 随后通过qPCR或高通量测序可实现多蛋白复合组成的精确鉴定与定量分析. 应用该技术, 研究团队系统解析了乳腺癌细胞系中34个关键肿瘤相关蛋白的相互作用网络, 发现肿瘤细胞内高阶蛋白复合体的多样性与丰度显著高于正常细胞, 提示这类复合体具有成为新型肿瘤生物标志物的潜力.
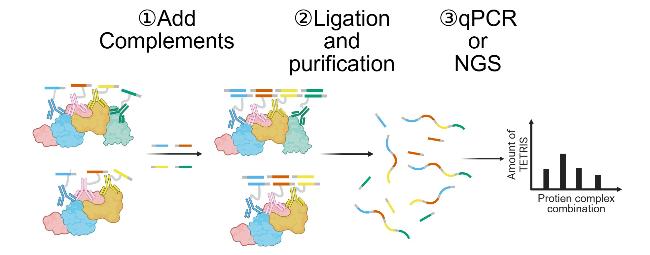
相较于已有的PPI检测方法, TETRIS技术能够实现多蛋白复合物(≥3个蛋白)的原位编码与并行检测, 突破了传统方法单次无法检测3个以上靶标蛋白的技术瓶颈. 未来, 可进一步结合机器学习算法对复杂PPI网络进行深入解析, 不断挖掘TETRIS在基础研究与精准医学诊断中的多元化应用潜力.
4 信号读出技术
邻近依赖性核酸自组装技术通过将PPI事件转化为特异性DNA序列, 并结合RCA、HCR或PCR等核酸扩增体系, 实现信号放大. 在此技术框架下, 如何将DNA序列信息高效转化为可定量解析的物理或化学信号, 成为技术落地的核心挑战, 其关键在于构建适配不同应用场景的信号读出模块. 目前, 核酸序列信息的读出方式主要包括荧光和电化学[42]等. 荧光读出的核心原理在于将带有荧光标记的DNA分子与因PPI而产生的特异性核酸产物进行杂交, 从而将核酸序列信息转化为光学信号, 其检测通常依赖具备高灵敏成像能力的荧光显微镜或流式细胞仪等光学设备. 此外, 高通量测序技术因其数字化特性和多路并行解析能力正成为PPI检测的有力手段, 其核心原理则是对DNA序列直接进行碱基识别, 将其精确转化为可分析的数字化字符信息(A、T、C、G). 本节将重点评述荧光成像与测序技术在 PPI 检测中的最新进展, 并深入探讨各自的技术原理与应用优势.
4.1 基于荧光的信号读出
依托核酸化学合成技术的不断发展以及核酸修饰的便捷性, 荧光标记策略已成为最成熟的核酸检测读出方法. 目前, 细胞水平的PPI检测主要依赖两大光学平台: 共聚焦显微成像与成像流式细胞术, 二者分别在空间分辨率与检测通量上满足不同应用需求.
激光扫描共聚焦显微镜(Laser Scanning Confocal Microscopy, LSCM)通过双针孔共轭光学系统实现微米级空间分辨率, 可通过光学切片能力消除离焦平面荧光干扰, 并支持PPI的亚细胞器水平空间定位分析. 2018年, Berndt-Paetz团队[43]将PLA与LSCM三维建模相结合, 成功绘制了膀胱平滑肌细胞中M3毒蕈碱受体与不同G蛋白亚型的动态互作图谱, 揭示了β-arrestin介导的受体内吞伴随Gαq/11亚型解离效率下降的新型G蛋白偶联受体信号脱敏机制.
成像流式细胞术(Imaging Flow Cytometry, IFC)通过整合流式细胞仪的高通量优势与显微成像的空间信息捕获能力, 是单细胞多参数PPI分析的新工具. 2017年, Abramson等[44]开发的邻近连接成像流式细胞术(Proximity Ligation Imaging Cytometry, PLIC)可用于稀有细胞中PPI的定量检测. 应用该技术, 研究人员发现大约25%的髓质胸腺上皮细胞中存在Aire蛋白与Sirt1蛋白的相互作用.
随着荧光检测平台的迅猛发展, 基于荧光的核酸序列信息读取将更为精准、便捷. 此外, 人工智能技术的介入正在重塑荧光信号的定量分析范式. 以空间蛋白质相互作用图神经网络(spPPI-GNN)[33]为例, 其通过Delaunay三角剖分构建细胞亚区室互作网络, 在非小细胞肺癌细胞的奥希替尼治疗耐药模型中, 识别到Sox2/Oct4互作丰度与药物响应呈负相关. 此类技术的开发标志着PPI检测正在从定性描述向网络化解析转型.
综上, 荧光检测技术已由早期的单细胞定性成像, 迭代至定量分析, 直至PPI网络图谱的绘制. 随着超分辨显微技术突破衍射极限, 以及深度学习在微弱信号提取中的应用, 基于荧光的信号读出方式将在临床诊断、高内涵药物筛选等领域展现更加重要的价值.
4.2 基于高通量测序的信号读出
基于高通量测序平台的信号读出方式在PPI的高通量解析中展现出革命性优势. 传统的显微成像技术因荧光淬灭、光谱串扰以及衍射极限等因素[45]难以高通量测定PPI; 而依托于高通量测序平台的信号读出方式, 突破了光学成像的物理屏障. 在引入了唯一分子标识符(Unique Molecular Identifier, UMI)后, 不仅可实现单分子精确追踪, 还能对PPI组分进行高保真识别.
2022年, Tay团队[46]通过整合邻近连接与单细胞测序技术, 成功开发出具有PPI解析能力的邻近测序(Prox-seq)平台, 在单细胞水平上实现了PPI网络与转录组的关联分析(图6). 借助基于随机事件概率模型的动态校正算法, Prox-seq能够区分稳定复合物信号与瞬时邻近噪声, 有效克服了传统荧光成像中因光学衍射极限、光谱重叠等因素造成假阳性结果的限制. 在对人外周血单核细胞的系统性研究中, Prox-seq首次鉴定出CD8/CD9跨膜复合体, 并通过单细胞转录组分析揭示其特异性呈现于幼稚CD8⁺T细胞亚群, 为T细胞分化调控机制研究提供了新的分子标志. 同样, TETRIS平台[41]亦通过基于UMI与高通量测序的解析技术对72例乳腺癌活检样本进行了高阶蛋白复合物的精准定量解析, 并利用组合数学模型将测序结果精确映射至原始复合物的组成与丰度, 体现了高通量测序在PPI信息分析中的独特优势.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
从光学成像到高通量测序的信号读出方式演进, 标志着PPI研究进入数字化定量时代, 为高分辨率、可溯源的动态PPI网络图谱绘制奠定了坚实的方法学基础. 这种兼具单分子精度与多组学纬度的创新技术体系, 为绘制时空分辨PPI图谱提供了有力工具.
5 总结与展望
邻近依赖性核酸自组装技术通过将PPI事件转化为特异性核酸信号, 并结合信号放大策略与高信噪比读出系统, 在PPI检测领域展现出独特优势. 在灵敏度方面, TETRIS平台通过基于PCR的核酸扩增技术达到了阿摩尔级别的检测限, 能够捕获样本中极其稀有的PPI事件; 在通量方面, iseqPLA技术通过迭代式探针擦除策略与多轮成像成功解析了非小细胞肺癌细胞在奥希替尼治疗前后多种PPI的动态变化; 在样本兼容性上, 已实现多种临床组织样本中PPI的精准检测, 并突破了基于质谱的PPI检测对样本量的苛刻要求.
与其他主流PPI检测方法相比, 如邻近依赖的生物素标记鉴定(Proximity-dependent Biotin Identification, BioID)[47]、双分子荧光互补(Bimolecular Fluorescence Complementation, BiFC)[14]等, 邻近依赖性核酸自组装技术聚焦于预设蛋白靶点的高维解析领域, 具有单细胞检测灵敏度和同时检测上百PPI的通量. BioID等邻近标记技术通过酶促生物素标记实现对靶标蛋白全局性互作的筛选, 特别适用于发现靶标蛋白的未知PPI, 但其依赖于大量细胞的同时标记, 检测灵敏度较低, 难以实现单细胞低丰度互作蛋白的精准检测. BiFC能够在活细胞中可视化PPI事件, 同时提供原位空间信息, 适用于PPI的动态追踪研究, 但其受限于遗传操作依赖性, 可能导致靶标蛋白的亚细胞定位异常或功能受损, 并且BiFC的检测灵敏度和通量均较低.
尽管已广泛用于科学研究领域, 邻近依赖性核酸自组装技术的临床应用仍面临多种挑战: 第一, 探针质量依赖性导致检测特异性受限, 抗体/核酸适体的亲和力参数与交叉反应率直接影响检测准确性; 第二, 靶标预判悖论, 基于已知表位设计的探针体系难以捕获新型PPI; 第三, 临床转化障碍, 多步骤操作流程带来的技术复杂性与设备依赖性制约其在即时检测场景的应用; 第四, 检测平台的物理极限, 荧光信号系统虽能获取PPI原位信息, 但受限于光学衍射极限难以实现单分子级成像, 而测序平台虽可实现单分子检测却无法溯源PPI的空间信息.
展望未来, 邻近依赖性核酸自组装技术亟需与计算生物学、人工智能、微流控芯片等跨学科技术手段深度融合, 以进一步突破灵敏度与应用边界. 在探针依赖性问题上, 可借助AlphaFold2等结构预测工具与深度学习平台[48], 基于靶标三维结构从头设计高亲和力抗体探针库. 曹龙兴与谢琦团队[49]通过蛋白质从头设计, 成功构建靶向胶质母细胞瘤EGFR和CD276的全新结合蛋白, 在CAR-T疗法中展现出优于传统单链抗体的实体瘤治疗效果, 为人工智能优化探针特异性提供了实证支持. 结合计算生物学与人工智能技术(如吴迪团队的KGF-GNN模型[50]), 可高效预测潜在的未知PPI, 进而通过邻近依赖性核酸自组装技术进行实验验证. 此类“计算预测→实验验证”闭环策略, 能将靶标预判悖论转化为技术优势, 充分发挥邻近依赖性核酸自组装技术高通量、高灵敏度特性. 针对临床转化障碍, 可以构建标准化全流程体系, 通过微流控技术[51-52]构建自动化反应模块; 整合纳米孔测序[53-54]实现实时信号读出; 引入人工智能算法自动解析数据, 大幅压缩操作时间, 建立“自动化反应→信号高效转换→智能读取”的一体化平台, 该流程可匹配即时检测需求, 推动技术临床落地. 通过上述跨学科创新, 邻近依赖性核酸自组装技术有望在精准医学诊断和新药开发领域掀起范式性变革.
Cheng, F.