


1 Introduction
Nucleic acids, as the core carriers of genetic information for life, have dynamic conformational diversity and complexity far beyond the traditionally known double helix model. In addition to classical B-form deoxyribonucleic acid (B-DNA), non-classical structures such as G-quadru- plexes (G4s), i-motifs, triplexes, and cyclic nucleic acids are formed through π-π stacking, hydrogen bonding networks, and cation coordination, and play an indispensable role in various key biological processes such as telomere protection, gene transcription regulation, and epigenetic modification.[1-3] For example, G4s consist of guanine-rich sequences that form a tetrameric plane through Hoogsteen hydrogen bonding, and their dynamic folding properties are widespread in gene promoters, telomeres, and oncogenes, which are involved in tumorigenesis by regulating transcription factor binding or replication fork stalling.[4-7] Whereas i-motif, as a tetrasome formed in cytosine- enriched regions, its acid-base responsiveness gives it a unique function in cell cycle regulation.[8] Abnormalities in the dynamic equilibrium of these structures within the cells are often closely associated with the onset and progression of a variety of malignant diseases, including, but not limited to, tumors, neurodegenerative diseases, and metabolic disorders.[9-11] As a result, they have become important diagnostic targets and potential targets for pharmacological intervention in precise medicine.[12-15]
Traditional techniques in nucleic acid detection such as gel electrophoresis, nuclear magnetic resonance (NMR)[16] and circular dichroism (CD)[17] can realize the qualitative analysis of nucleic acid structure, but they have limited spatial resolution (usually greater than 100 nm), destructive sample processing and lack of real-time dynamic monitoring ability. In recent years, fluorescence probes have gradually become a cutting-edge tool in detection of non-classi- cal nucleic acid structure due to their excellent sensitivity, fast response, good biocompatibility, excellent real-time dynamic monitoring ability, and programmable modifiability.[18-22] For example, small-molecule fluorescent dyes enable rapid imaging by directly labeling nucleic acid grooves; macromolecular fluorescent materials, such as conjugated polymers, take advantage of π-π stacking to bind to nucleic acids and enhance the resonance energy transfer efficiency;[23] quantum dots can be used for multicolor labeling and high-sensitivity signal amplification by virtue of their size-tunable luminescence,[24-27] and upconverting nanoparticles (UCNPs), which can penetrate into deep tissues because of near-infrared excitation, enable nondestructive real-time monitoring of the presence of nucleic acids in live bodies and in living tissues[28,29]. Therefore, by designing fluorescence probes under these principles, they are expected to specifically bind to the grooves or terminal planes of the target nucleic acid structure, and realize the amplification and output of the detection signals by utilizing conformational changes or resonance energy transfer me- chanisms.
In this review, we systematically summarize the fluorescence probes applied for the detection of G4s and other non-classical nucleic acid structures (such as i-motif, Triplex, and cyclic nucleic acids), revealing the unique advantages of different probe types in structure recognition and signal transduction. By detailing the design principles, detection mechanisms, and application scenarios of fluorescence probes such as traditional small molecular fluorophores and aggregation-induced emission luminogens (AIEgens) for non-classical nucleic acid structure detection, we reveal their potential value in disease diagnosis and precise medicine. By systematically optimizing the structure-property relationship and mechanism of target-recog- nition and signal output, we expect to build a highly selective, highly sensitive, and multi-channel responsive platform, which will provide an innovative tool for the analysis of life regulatory networks and the development of next- generation theranostic technologies.
2 G-quadruplexes (G4s)-response fluore- scence probes
G4s are π-π stacked tetramer structures formed by four guanine residues bonded by Hoogsteen hydrogen bonds. G4s have unique topological polymorphisms, which can be categorized into parallel, antiparallel, and mixed conformations, with parallel conformations dominating due to low steric hindrance and high stability.[30] Its formation is regulated by multiple factors, including nucleic acid sequence length, base arrangement, cation type, solution viscosity and pH. For instance, a low pH environment can promote the stable conformation formation.[31] The biological functions of G4s are closely related to their role as protein- binding targets, and more than 20 proteins regulate G4s dynamic homeostasis through specific adhesion.[32-34] G4s abnormalities are closely associated with cancer, diabetes and neurodegenerative diseases, such as c-MYC promoter G4s overexpression to activate proto-oncogenes, and telomere G4s dysregulation to trigger chromosomal instabi- lity.[35] Therefore, G4s are considered important targets for disease diagnosis and treatment.[36-38]
The detection of G4s mainly relies on their unique topology and chemical properties. The detection mechanism consists of four main modes of action: (1) Electrostatic adsorption: positively charged probes bind to the phosphate groups in the G4 nucleic acid structure through Coulombic forces; (2) Conformational embedding: rigid aromatic com- pounds are inserted into the G4 groove and π-π stacking interactions occur; (3) End stacking: planar molecular probes bind tetrameric ends via side chain π-π interactions; (4) intertetramer insertion: amphiphilic molecules are inserted into adjacent tetramer gaps to promote interchain alignment and amplify signals. The development of fluorescence probes for G4s usually involves the design of different binding sites for the different binding modes mentioned above.
2.1 Elastine dyes
Essence dyes in molecular fluorescence probes (especially anatase dyes) occupy an important position in the research field of biosensing as well as disease diagnosis and treatment due to their good optical properties and high biocompatibility.[39] The basic structure of Anna dyes usually consists of two nitrogen-containing heterocycles connected by an unsaturated poly(hypromellitic) bridge (Figure 1a).[40] These two nitrogen atoms are the components of the heterocyclic nucleus, linked by chains of multiple hypomethyls. Heterocycles can be classified into various types, such as quinoline, thiazole, imidazole and the corresponding benzene systems. The length of the poly(hypomethyl) bridge and the terminal chromophores directly control the absorption and emission peaks of the dye. Many Anna dyes can interact specifically with nucleic acids and exhibit specific photophysical signaling response signals, which are widely developed for applications such as nucleic acid imaging and cancer diagnosis.[41]
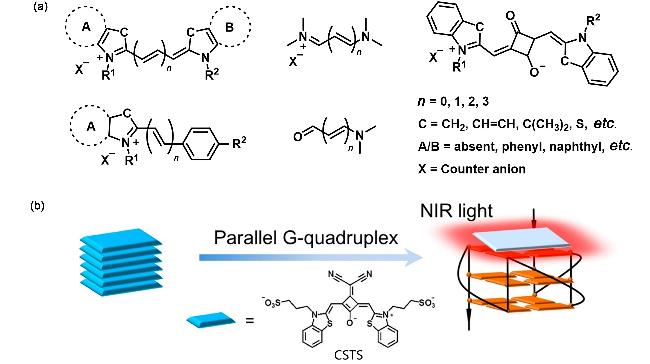
Squaraines are a class of long-wavelength anthocyanine dyes with a central electron-deficient quaternary ring nucleus connected by an electron-rich aromatic portion (Figure 1a). These dyes exhibit strong absorption and fluorescence emission in the red to NIR region. As shown in Figure 1b, Shangguanʼs group synthesized a squaraines dye, CSTS, which binds to G4s via an end-stacking model on the G4 surface, and fluorescence intensified significantly when interacting with parallel G4s in particular, but did not interact significantly with other DNA structures.[42] The high selectivity of CSTS for parallel G4s has been attributed to the bis(cyano-methylene) functionalized squaraine dyes with a V-shaped rigid planar π-scaffold. CSTS exists mainly in aggregated form in water, but is converted to monomeric form upon interaction with parallel G4s, which turns on the fluorescence.
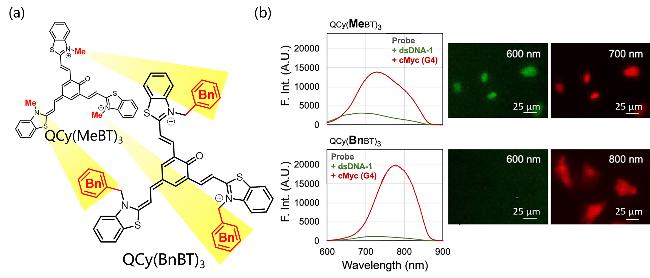
In addition, Sakamotoʼs group developed a one-donor-three-acceptor (D3A)-type ternary quinone-cyanine dye QCy(MeBT)3 (Figure 2a) with an N-methylbenzothiazole cation as an acceptor, which is capable of binding double-stranded DNA (dsDNA) and G4s with fluorescence switching properties in different colors (binding to dsDNA and G4s emits, respectively, a fluorescent signals at 600 and 700 nm wavelengths).[43] In addition, the team found that the two N-methylbenzothiazole cationic portions of QCy- (MeBT)3 were suitable for binding to the minor grooves of the A- and T-base-rich regions of dsDNA, whereas the introduction of bulky functional groups could effectively inhibit the ligand-dsDNA interaction. Thus, QCy(BnBT)3, obtained after the introduction of the benzyl-substituted methyl group, eliminates interaction with dsDNA but retains its terminal structure for stacking with the top plane of G4s. Upon contact with G4s, the fluorescence intensity of QCy(BnBT)3 was enhanced by at least 500-fold, achieving highly specific and sensitive recognition of G4s (Figure 2b).
Liʼs group developed a supramolecular assembly of the anionic elastin dye SCY-5 (Figure 3a) that can interact with G4s and dissociate into monomers [44]. The movement of the C—C bond is thus effectively inhibited, leading to a rapid and dramatic increase in fluorescence emission (Figure 3b). SCY-5 only fluorescently lights up parallel G4s but not antiparallel and hybridized G4s (Figure 3c). This specificity for parallel G4s stems from the charge and structure of SCY-5. First, SCY-5 has two negatively charged propylsulfonic acid groups, which strongly repel negatively charged nucleic acids and prevent SCY-5 from binding to single-stranded DNA (ssDNA) and dsDNA. Secondly, the G4 terminal planes of antiparallel and hybrid G4s are surrounded by different rings, which create stronger steric hindrance and affect the binding of SCY-5 to the G4 terminal planes, and only the parallel G4s have less site resistance, which allows SCY-5 to stack on the terminal planes. SCY-5 was validated to be able to successfully quantify ribonucleic acid (RNA) G4s extracted from venous bloods of patients with pancreatic, breast, and hepatocellular carcinomas.
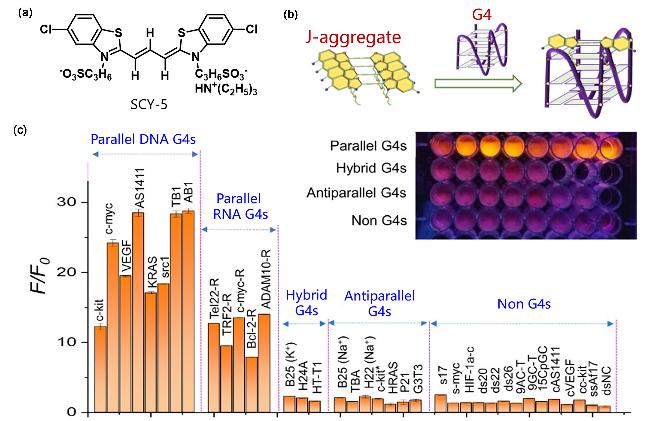
Figure 3 (a) Structural formula of SCY-5; (b) Luminescence phenomenon of SCY-5 inducing the disassembly of parallel G4s upon binding; (c) Fluorescence intensity (F/F0) of SCY-5 at 590 nm in the presence of different nucleic acid modelsReproduced from [44]. Copy right 2024 American Chemical Society. |
Zhangʼs group[45] synthesized a novel switching fluore-scent elastomeric dye SN-Cy5-S based on a benzothiophene rotor for selective recognition of G4 structures in mitochondria. The probe is based on SN-Cy5, which introduces flexible mesoaromatic heterocyclic substituents on the polymethyl chain, weakening the chimeric interaction with dsDNA while also providing a larger surface area for π-π stacking with G4s. As shown in Figure 4a, the entire SN-Cy5-S molecule can form a nearly planar “Y” shape by end-stacking on the top plane of the 5' end of G4s. At this point, SN-Cy5-S, which is self-quenched in solution by H-aggregation, undergoes depolymerization, and the twist- ing of the C—C bond between polymethyl chain and m- benzothiophene is restricted, triggering strong fluorescence enhancement with a detection limit as low as 1.51 nmol/L (Figure 4b). The researchers applied SN-Cy5-S for targeted imaging of G4s in mitochondria with staining concentrations as low as 0.5 μmol/L and response times within 5 min (Figure 4c).
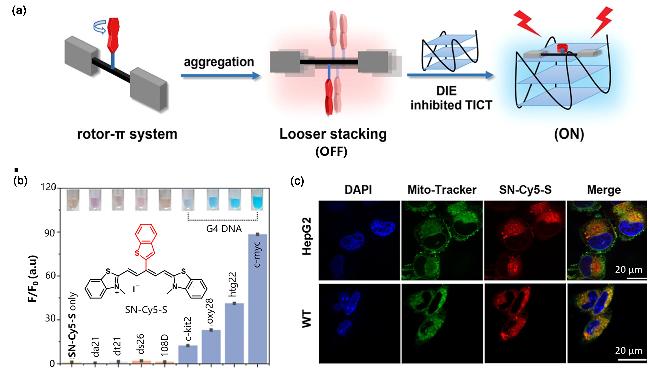
Figure 4 (a) Combination of SN-Cy5-S monomer with G4s inhibited twisted intramolecular charge transfer (TICT) and turn-on fluorescence; (b) Fluorescence enhancement (F/F0) of SN-Cy5-S after interaction with different nucleic acids; (c) Confocal fluorescence image of SN-Cy5-S mitochondrial co-localizationReproduced from [45]. Copy right 2023 American Chemical Society. |
2.2 Thiazole orange dyes
Thiazole orange (TO) dyes belong to the cyanine dye, which are asymmetric anthocyanin dyes with unique fluorescent properties. TO is often used for the staining and quantification of reticulocytes, where it produces a substantial fluorescence enhancement and high quantum yields upon binding to nucleic acids, especially RNA.[46]
Kotlyarʼs group[47] found that TO was able to form very stable complexes with triplexes (Triplex) and G4s that did not dissociate during column separation and gel electro-phoresis and thus could be used for Triplex and G4s detection, whereas dsDNA could not form stable complexes with TO (Figure 5). More importantly, the combination of TO with Triplex and G4s resulted in a significant (>1000-fold) increase in fluorescence intensity. Because the absorption spectra were red-shifted (ca. 14 nm) and blue-shifted (ca. 15 nm) upon binding to Triplex and G4s, respectively, TO could be used to differentiate between Triplex and G4s. CD spectroscopy showed that TO also induced different chiral signals on Triplex and G4s, further demonstrating its ability to differentiate between Triplex and G4s.
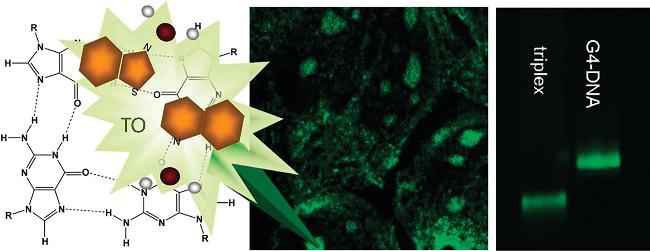
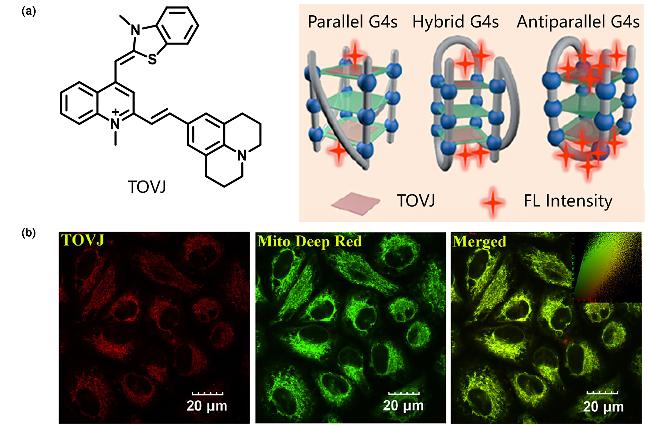
An another TO-derived probe, TOVJ was synthesized by Luʼs group[48] via further introducing a Julolidine unit into the TO molecule, which significantly improved its fluorescence properties. TOVJ was much more selective for G4s than other DNA topologies (such as ssDNA/dsDNA). In particular, it recognized antiparallel G4s much more effectively. As shown in Figure 6a, the enhanced selectivity of TOVJ over G4s is due to the ability of the side chain of TOVJ to interact with the grooves and ring regions of G4s. TOVJ showed significant fluorescence enhancement when interacting with G4s, especially antiparallel G4s, with up to 2742-fold enhancement at emission maximum locating in the near-infrared region (694 nm). The detection limits of TOVJ for G4s were as low as 164 pmol/L (Hum24) and 231 pmol/L (22AG Na+). In addition, TOVJ was able to enter living cells and target mitochondria, suggesting its potential in intracellular G4s imaging (Figure 6b).
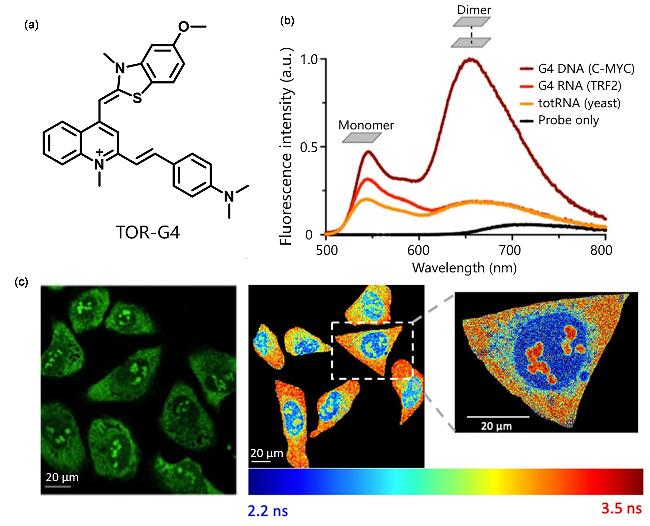
A new thiazole orange derivative TOR-G4 was synthe- sized by Kuimovaʼs group.[49] As shown in Figure 7a, the researchers modified the phenylstyryl unit on TO, extending the degree of electron conjugation of its original motif, thereby enhancing the π-stacking potential with G4s (Figure 7b). Methoxy and dimethylamine groups are introduced to both ends of the probe, and the positively charged nitrogen is sandwiched in the molecular center to enhance its two- photon excitation for imaging within cells and tissues. As the combination with G4s can significantly increase the fluorescence lifetime of TOR-G4, the fluorescence lifetime imaging microscopy (FLIM) technique was thus successfully used to monitor the dynamics of intracellular G4 (Figure 7c).
2.3 Thioflavin dyes
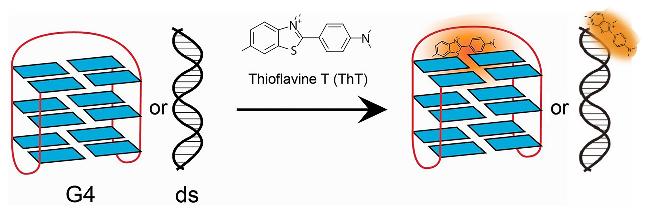
Thioflavin T (ThT) is a cationic benzothiazole dye that is cell membrane permeable and is commonly used to identify amyloid fibrils in tissue samples. Thioflavin T generally has an optimal absorption wavelength around 440 nm, emits yellow-green fluorescence, and has an optimal emission wavelength near 490 nm. However, upon binding to the target, its fluorescence excitation and emission wavelengths are shifted. There are many reports that thioflavin T and its derivatives have specific recognition ability for G4s. As early as 2014, Mergnyʼs team verified that the fluorescence of ThT was significantly enhanced in the presence of most G4s forming sequences, while the fluorescence enhance-ment was limited in the presence of dsDNA and ssDNA sequences (Figure 8).[50]
Tangʼs group[51] synthesized a small-molecule ThT analog, IMT, which interacts with G4s by simply stacking in the 3'-G4 terminal plane (Figure 9a). It is virtually non- fluorescent in an aqueous environment, but exhibits significant fluorescence enhancement when interacting with G4s structures (Figure 9b). This feature enables IMT to stain G4s in vitro and in living cells with a high signal-to-noise ratio (Figure 9c).
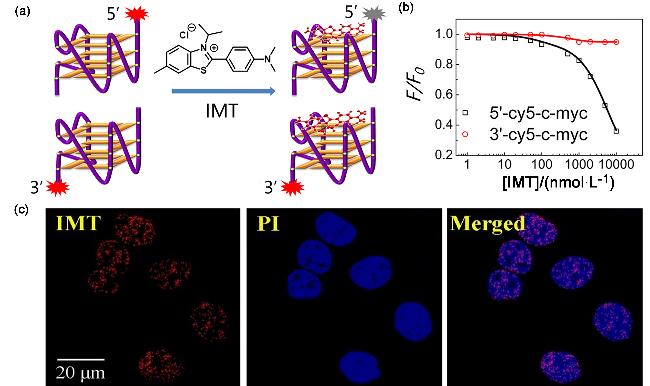
Figure 9 (a) Schematic diagram of the selective binding of IMT and G4s; (b) Relationship between F/F0 after the combination of IMT and c-kit and the concentration of IMT; (c) Confocal imaging of Hela cells stained and fixed with IMT and PIReproduced from [51]. Copy right 2018 Oxford University Press. |
A benzothiazole-based fluorescence probe ThT-NE was developed by Nieʼs group for RNA G4s imaging in living cells.[52] The presence of a certain twist angle between the methylbenzothiazole and N,N-diethylimine portions of the molecule results in a twisted intramolecular charge transfer (TICT), and thus ThT-NE alone is fluorescence quenched. When it binds to G4s, the π-π stacking interaction between ThT-NE and the terminal G4 planes of G4s significantly restricts the intramolecular rotation of ThT-NE, thus turning on fluorescence (Figure 10a). The team used the probe for direct intracellular visualization of G4s sequences (CG2a) in hepatitis C virus (HCV) RNA, as shown in Figure 10b.
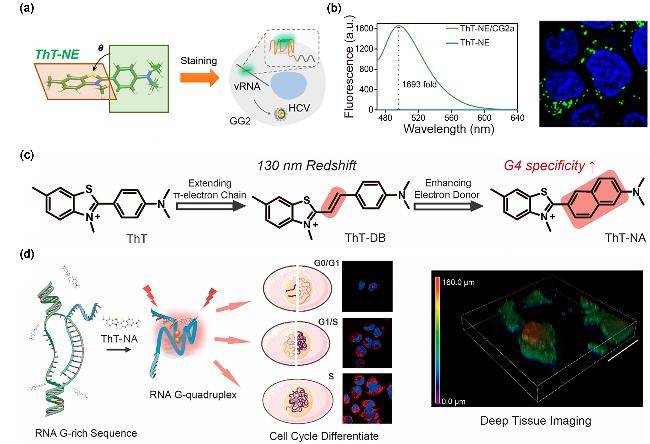
Figure 10 (a) Schematic diagram of the principle of detecting HCV vRNA using G4s probe ThT-NE; (b) Fluorescence spectra of ThT-NE in the presence of CG2a and direct intracellular visualization of CG2a by this probe; (c) Molecular structure, probe properties and modification of ThT; (d) Differentiation of cell cycle by ThT-NA through RNA G4 formation signal(a, b) Reproduced from [52], Copy right 2019 American Chemical Society.; (c, d) Reproduced from [53], Copy right 2022 American Chemical Society. |
ThT-NE fluorescence was enhanced more than 1693-fold in the presence of CG2a, demonstrating high selectivity and sensitivity. In addition, Nieʼs team synthesized another ThT-NA for detecting and imaging G4s structures in live cells and tissue samples[53]. The molecule binds to G4s by a mechanism similar to that of ThT-NE, and by enhancing the electron donor and optimizing the molecular structure, ThT- NA exhibits excellent photophysical properties (Figure 10c), including red light emission (610 nm), large Stokes shift (>100 nm), and high selectivity (1600-fold fluorescence turn-on ratio) for G4s. As shown in Figure 10d, ThT-NA was able to reveal the distribution of endogenous RNA G4s in living cells and differentiate the cell cycle by monitoring the folding of RNA G4s.
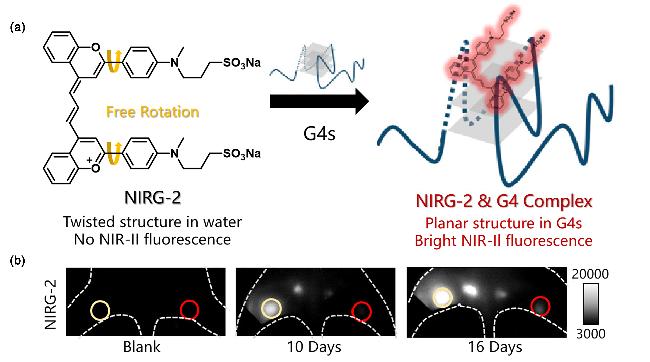
Apart from these visible range emissive probes, a NIR-II fluorescence probe, NIRG-2, based on the thioflavin T analog anthocyanin (N-Cy), was designed by Zhangʼs group for specifically in vivo G4s detection.[54] As shown in Figure 11a, the sulfonic acid moiety in the NIRG-2 molecule is able to form stable hydrogen bonds with specific nucleotides (such as DG-15) in the structure of G4s. At the same time, the NIRG-2 molecule forms multiple π-π stacking interactions with guanine bases in G4s through its benzene ring structure. These interactions further stabilized the probe complex with G4s and effectively suppressed the twisted intramolecular charge transfer (TICT), which enabled highly selective detection of G4s. NIRG-2 emits at 940 nm with a large Stokes shift of 90 nm, showing up to 47-fold fluorescence enhancement and a tissue imaging depth of 5 mm with excellent photophysical properties. Using NIRG-2, the research team successfully achieved high-contrast imaging of tumor metastasis and evaluation of the therapeutic effects of cancer lymphatic metastasis (Figure 11b).
2.4 Other small molecule dyes
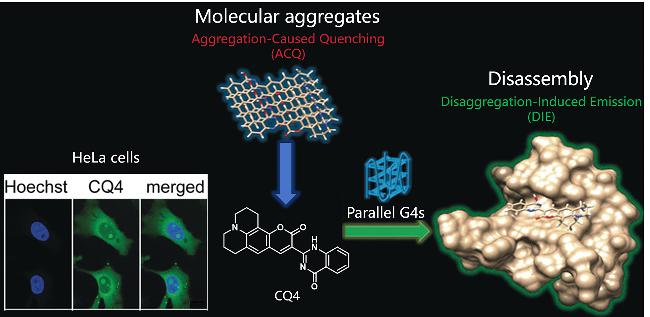
Sabouriʼs group[55] synthesized a dye CQ4 based on coumarin-quinazoline conjugates, which forms molecular aggregates in solution with aggregation-caused quenching (ACQ) characteristics. In the presence of parallel G4s, CQ4 binds to the terminal planes of the G4s structure via π-stacking, leading to the breakdown of the molecular aggregates into a highly emissive monomeric state, which turns on the fluorescence signal (Figure 12). On the contrary, antiparallel G4s and non-G4s structures could not completely replace the aggregated state of CQ4, and the fluorescence recovery was poor. The binding constants of CQ4 to parallel G4s structures were three orders of magnitude higher than those of non-parallel G4s structures, suggesting that CQ4 was highly selective for parallel G4s structures. The detection limit of CQ4 to c-MYC G4s was up to (83.5±4.2) nmol/L with linear range from 0 μmol/L to 1.40 μmol/L. In fixed HeLa cells CQ4 preferentially stained the nucleolus region, and in live cell, CQ4 co-localized with the mitochondrial network, indicating its intracellular targetability.
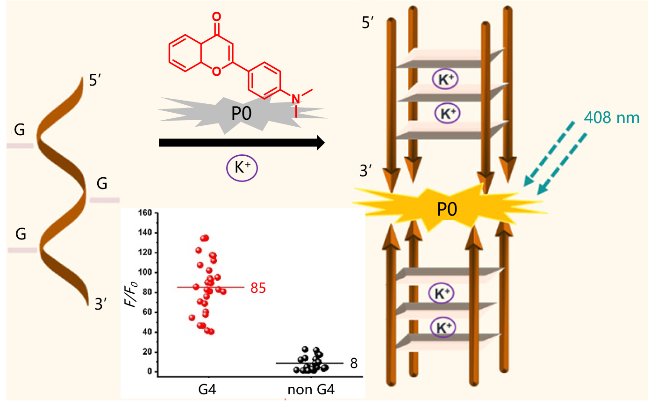
Another fluorescence probe for G4s was designed based on flavonoid P0, which efficiently probes G4s confoma- tions without disturbing their folding dynamics (Figure 13).[56] In the presence of G4s, the fluorescence intensity of flavonoid P0 was significantly enhanced to 85-fold, while it was only 8-fold when interacting with other DNA stru-ctures, indicating its high selectivity in G4s detection.
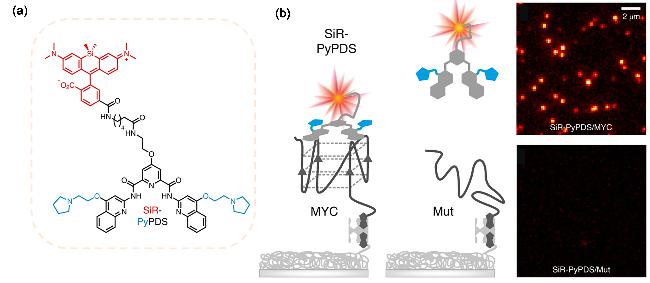
Based on the known structure of the G4s ligand PyPDS, Balasubramanianʼs group[57] designed a novel fluorescence probe called SiR-PyPDS for single-molecule and real-time detection of individual G4s structures in living cells. SiR- PyPDS is a connection of far-infrared fluorophore silrothramine (SiR) to an analogue of the known G4s ligand Pyridostatin (PyPDS) (Figure 14a), which specifically binds G4s by forming hydrogen bonds and hydrophobic interactions with guanine bases in the G4 structure. As shown in Figure 14b, SiR-PyPDS was able to detect an average of 867 long-lived binding events at a low concentration (250 pmol/L), whereas the binding events for the control probe SiR-iPyPDS (a non-G4s ligand) were reduced to 66 events, and the probe was capable of specifically bind G4s at low nanomolar concentrations, avoiding overall interference with G4 folding kinetics. In U2OS cells, SiR-PyPDS was able to detect binding events of individual G4s with a short duration of the binding event (6.6±0.5 s), suggesting that G4s are in a dynamic folding state within the cell. In addition, the formation of G4s is verified to be closely related to the cell cycle and is regulated by transcription and replication activities, providing a powerful tool to decode the specific biological functions of G4s in the human genome.
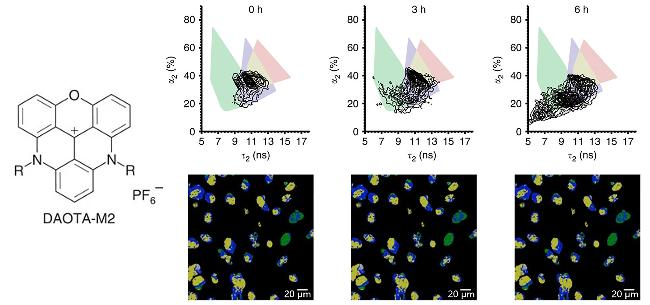
Except these intensity-response probe, fluorescence lifetime was another measurement parameter in the detection of G4s. Vilarʼs group[58] designed a new triangulenium derivative DAOTA-M2 with significant changes in fluorescence lifetime when interacting with different nucleic acid topologies. In particular, DAOTA-M2 shows a longer fluorescence lifetime upon binding to G4s compared with ssDNA and dsDNA. As shown in Figure 15, leveraging fluorescence lifetime imaging microscopy (FLIM) technique, researchers were able to observe the interaction of small molecules with G4s in real-time manner in living cells.
2.5 Bioactive substance-based probes
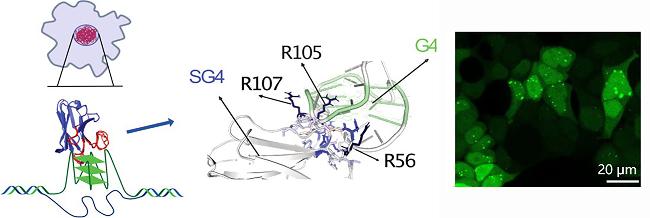
Leveraging the specific interaction with antigen, binding-emission fused nanoantibodies were also designed for G4s detection. Nanobodies are relatively small antibodies (about 12 to 15 kDa) whose antigen recognition is mediated by three structural domains of proteins called complementary decision regions (CDRs). The longest CDR3 is thought to play a central role in antibody-antigen interactions, enabling the recognition of epitopes not recognized by conventional antibodies. As shown in Figure 16, Balasubramanianʼs group[59] screened a novel camel heavy chain nanoantibody, SG4, specifically for the recognition and binding of human c-MYC G4 structure. SG4 exhibits sub-nanomolar levels of affinity for a wide range of folded G4 structures, and this specific affinity is derived from the arginine residue in CDR3. By fusing SG4 with green fluorescence protein (GFP), researchers were able to express and visualize the SG4-GFP-FLAG fusion protein in living cells. The expression of this fusion protein allowed researchers to localize G4s structures by directly observing G4s focal points in the nucleus through fluorescence microscopy, providing a new approach to further investigate the function of G4s in gene regulation and telomere biology.
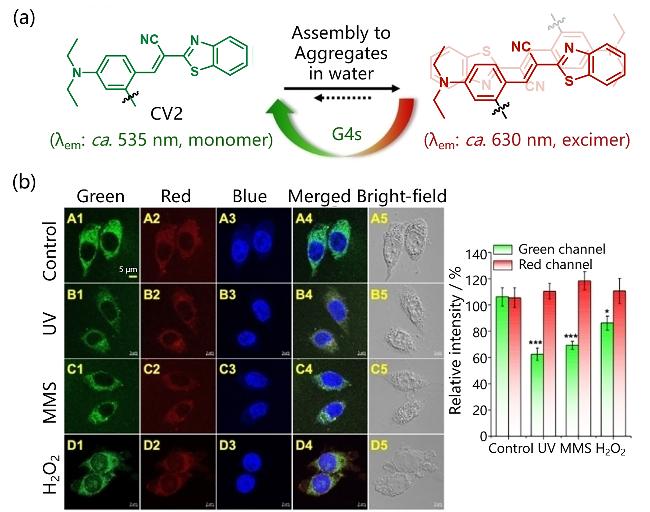
By binding a cyanoethylene dye to a recognition peptide sequence (l-Arg-l-Gly-glutaric acid), Kimʼs group[60] synthesized a self-assembled fluorescence nanoprobe, CV2, for selective imaging of parallel G4s structures in mitochon-drial DNA. As shown in Figure 17a, CV2 forms self-assembled nano-aggregates with red luminescence in water, but upon binding to G4s, it depolymerizes into rigid monomeric dyes, resulting in a significant enhancement of fluorescence. Highly selective and quantitative detection of c-MYC G4 for parallel G4s structures was realized with a good linear relationship in range of 0~0.5 μmol/L and a limit of detection of about 35 nmol/L. In addition, CV2 can be used to track the folding and unfolding state of the mitochondrial G4s structure in living cell, providing a valuable tool for further studies of mitochondrial oxidative stress and other mitochondrial DNA damage (Figure 17b).
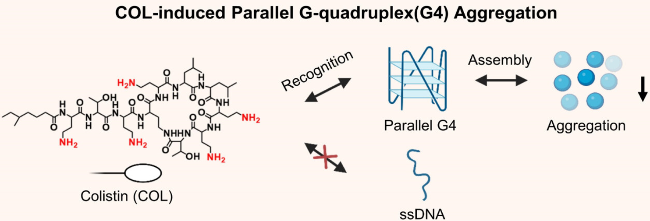
As a bacterial RNA G4s interfering agent, colistin (COL) is a cationic decapeptide previously used against drug- resistant bacteria. As shown in Figure 18, Zhouʼs team[61] proposed a new method called CoRP-seq based on COL, which was used to identify RNA G4s from HeLa cell lysates within 15 min without magnetic bead (MB) purification. Due to the electrostatic interactions between COL and G4s, as well as effective π-π stacking, COL can selectively induce the aggregation of G4s (especially parallel G4s) from nucleic acid mixtures. After G4s is assembled into aggregates in the presence of COL, the structure of G4s will not be denatured. At this point, the emission fragments such as N-methyl-mesoporphyrin IX (NMM) and ThT can be modified to achieve further fluorescence imaging of G4s.
2.6 Aggregation-induced emission probes
Fluorescence imaging technology plays an important role in the field of biomedical detection due to its advantages of non-invasiveness, high sensitivity, and high spatiotemporal resolution. However, conventional fluorescence dyes are susceptible to phase separation from hydrophilic media in physiological buffers due to their hydrophobic aromatic ring structure and π-π coupled chromophores, which form aggregates through π-π stacking, leading to significant quenching of fluorescence emission. This aggregated fluorescence attenuation effect severely limits the detection sensitivity and reliability of the probe. In contrast, aggregation-induced emission materials (AIEgens) show almost no fluorescence emission in dilute solution, but exhibit significant fluorescence enhancement in high concentration or aggregated state.[62] This special optical behavior originates from the restricted intramolecular motion (RIM) mechanism. When AIEgens with propeller-like molecular structure are aggregated, the original freely rotating molecular rotor is restricted by steric hindrance, and the non-radiative energy dissipation pathway is effectively suppressed, thus efficiently directing the excitation energy to the radiative excursion process.[63-67] Compared with traditional planar organic fluorophores, AIEgens can spontaneously aggregate in hydrophilic media or achieve "light-up fluorescence" function by specific binding with analytes, showing excellent detection sensitivity and signal-to-noise ratio, and multiple advantages such as high quantum yield, excellent light stability, photobleaching resistance, and large Stokes shift.[68-76] Currently, many research groups have utilized AIEgens to provide innovative solutions for nucleic acid detection with high sensitivity, multi-targetability, and real-time operability.[77-83] In addition, the AIE principle can be applied to develop G4s fluorescence probes because G4s can always accommodate responsive ligands, which rigidify their conformations and restrict intramolecular motions, thereby, triggering a responsive fluorescence signal.
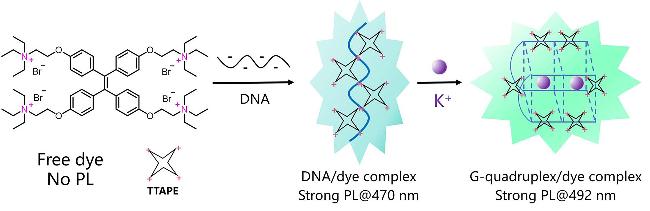
As the AIE research pioneer, Tangʼs group[84] developed a label-free DNA detection system, 1,1,2,2-tetrakis[4-(2- triethylammonioethoxy)phenyl]ethene tetrabromide (TTA- PE) that exhibited significant luminescence changes in G- rich DNA recognition via electrostatic attraction (Figure 19). First, when G-rich DNA strands (G1) were added to the TTAPE solution, the luminescence intensity increased significantly and showed a good linear relationship within a certain concentration range. The addition of K+ ions to the G1 solution induced the formation of G4s, and the luminescence intensity of TTAPE gradually recovered after the initial stage of decline, and finally reached a stable state, and the fluorescence spectra were red-shifted, which indicated that TTAPE could recognize G4s and monitor the process of the folding of G1 to form G4s in real time.
Reproduced from [84]. Copy right 2008 Wiley Online Library.
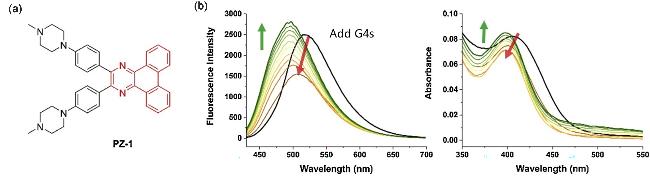
Huʼs group[85] chose the self-assembled luminophore dibenzoquinoline as the fluorescent core on which two piperazine-substituted phenyl rings were introduced to obtain the PZ-1 probe (Figure 20a). The introduction of these groups not only enhanced the π-π stacking interaction with G4s, but also provided additional electrostatic interactions between the positively charged amino group of G4s and the phosphate backbone. In addition, PZ-1 has a highly distorted conformation due to the spatial repulsion between the two benzene rings, which hinders intermolecular face- to-face stacking. In contrast, upon binding to G4s, the intramolecular rotation of the phenyl rings was restricted, subsequently leading to fluorescence enhancement. As shown in Figure 20b, a strong enhancement in fluorescence signal was displayed at 515 nm when PZ-1 was bound to c-MYC.
By combining TPE and a thiazole orange derivative (mTO) via the Knoevenagel reaction, Yuʼs group[86] synthesized a label-free fluorescence probe, TPE-mTO. As shown in Figure 21a, the expanded electron-deficient cation conjugation system and crescent-shaped scaffold of TPE-mTO enhanced its selectivity for the structure of G4s, and the positive charge of the mTO portion gave it mitochondria-targeting ability. TPE-mTO was rotationally blo- cked, leading to the turn-on fluorescence with a 38-fold enhanced intensity upon binding to G4s. It has high selectivity and sensitivity to G4s, and the detection limit is as low as 4.1 nmol/L. Live cell imaging demonstrated that TPE- mTO can specifically localize in mitochondria and clearly observe the structure of G4s (Figure 21b).
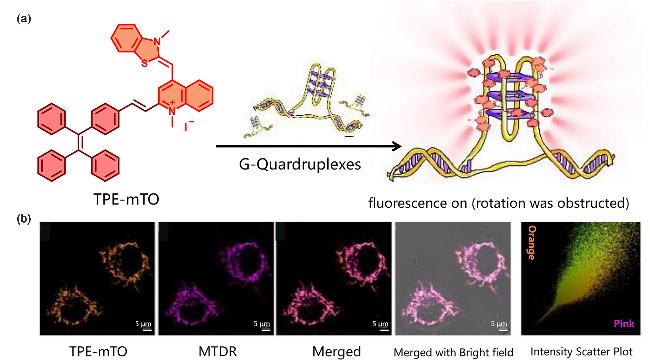
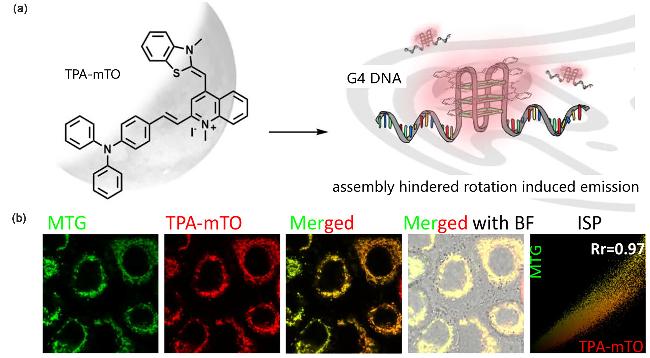
Another novel mitochondrial G4s probe, TPA-mTO, with near-infrared emission, ideal photostability, and high fluorescence contrast, was developed by the same group (Figure 21a).[87] TPA-mTO forms stable binding with G4s through multiple interaction modes such as π-π interactions, π-cation interactions, and hydrogen bonding. As shown in Figure 22b, in both live cells and fixed tissues, TPA-mTO probe was able to specifically bind to G4s structures in mitochondria with a 20-50-fold fluorescence enhancement without significant fluorescence enhancement (less than 5-fold fluorescence enhancement) with other nucleic acid structures (such as ssDNA and dsDNA), demonstrating its high selectivity for G4s. In addition, TPA-mTO also showed excellent photostability and long-term tracing ability in fixed cells, making it suitable for long-time imaging.
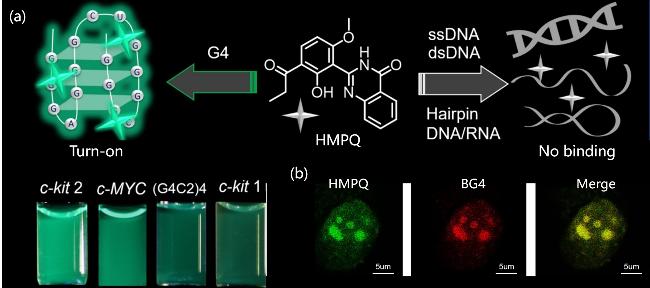
Qianʼs group[88] discovered a novel biological AIEgen 2- (2-hydroxy-6-methoxy-3-propionylphenyl)quinazolin-4- (3H)-one (HMPQ) from marine bacterial metabolites that can response to G4s. As shown in Figure 23a, the inherent biplane aromatic ring of HMPQ enables it to bind to G4s by π-π interactions, but not to conventional ssDNA, dsDNA, or hairpin nucleotides. HMPQ turns on fluorescence when it specifically binds to G4s. In particular, its fluorescence intensity is significantly enhanced by up to 300-fold upon interaction with c-kit2, and the fluorescence spectrum is red-shifted to 497 nm with a detection limit of 28.7 nmol/L. The overlap coefficient of HMPQ and the G4s structure- specific antibody BG4 was as high as 94.3% when detecting G4s in cells, demonstrating its high selectivity and sensitivity (Figure 23b).
The development of fluorescence probes has realized a multi-level leap from in vitro detection to in vivo imaging of G4s. Currently, probes based on π-π stacking, TICT effect and supramolecular assembly have achieved nanomolar- level sensitivity in extracellular detection; mitochondria-targeted probes combined with FLIM and other technologies have achieved dynamic tracking of G4s in living cells; and NIR-II probes have promoted in situ tumor visualization through deep tissue penetration. These technologies provide high spatial-temporal resolution for cancer diagnosis and treatment. In the future, the limits of single-molecule dynamic capture and deep-tissue monitoring can be resolved through introducing DNA origami probe arrays, light-activated femtosecond imaging, and microwave- emission probes. The cross-scale integration of technologies is expected to promote G4 probes to realize precise diagnosis and treatment in clinical scenarios such as liquid biopsy and early screening of neurodegenerative diseases, opening a new era of dynamic genomics research and translational medicine.
3 Development of fluorescence probes for other atypical nucleic acid structures
3.1 i-Motif sensing
The i-motif is a dynamic non-classical DNA secondary structure found primarily in cytosine-rich DNA sequences. Its core feature is formed by the binding of two parallel strands of dsDNA by intercalated C-C base pairs in antiparallel orientation, in which the cytosine bases are paired via Hoogsteen hydrogen bonds and depend on a protonated C4 position amino group for enhanced stability.[89] Unlike G4s that depend on purine base stacking, the stability of i-motif is highly dependent on the acidic environment (pH<6.5) and unfolds under neutral conditions due to deprotonation. This pH-responsiveness endows it with the property of reversible conformational changes.[90] i-Motif is widely involved in genome regulation and cellular homeostasis maintenance. It has been found in the promoter and telomere regions of human oncogenes, dynamically regulating gene transcriptional activity by binding transcription factors or blocking RNA polymerase.[91] In telomeres, i- motif inhibits telomerase activity, thereby limiting the ability of cancer cells to immortalize, leading to poor genomic stability and promoting apoptosis.[92] In addition, i-motif is involved in DNA replication, repair, and filament localization, suggesting its multiple regulatory roles in tumorigenesis and senescence. This structural specificity makes it a potential therapeutic target, especially in cancer therapy exhibiting a dual mechanism of blocking proto-oncogene expression and disrupting telomere maintenance, indicating the significance of the specific and sensitive detection of i-motif structure.
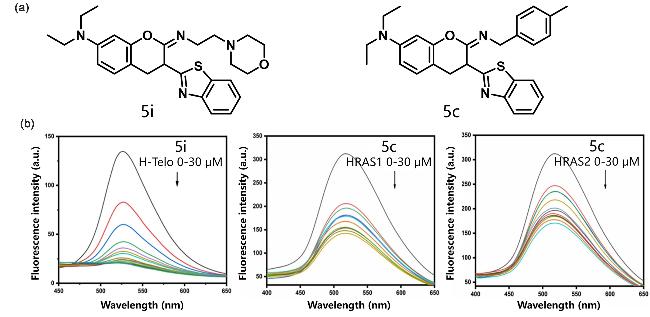
Bhowmikʼs group synthesized a series of benzothiazolyl coumarin derivative probes (5i, 5c) to explore their specific binding to i-motif (Figure 24a)[93]. By UV-Vis absorption and fluorescence spectroscopy, 5i was found to have significant affinity for H-Telo, 5c for HRAS1 and HRAS2 i-motif, with binding constants of 3.38×106, 9.07×105, and 9.35×105 L•mol-1, respectively, which were much higher than the binding constants for dsDNA (1.04×105 L•mol-1) (Figure 24b). The probes interacted with the bases in the i-motif by forming π-π stacking interactions between the unoccupied antibonding orbitals (π*) of the compounds and the bonding orbitals (π) of the DNA bases, resulting in enhanced fluorescence intensity and increased fluorescence lifetime, indicating their potential for the detection of i-motifs.
Another fluorescence probe, G59 was developed by Dingʼs group[94] for the specific identification and detection of the i-motif structure of the c-MYC gene promoter (Figure 25a). G59 combines carbazole and naphthylimide fluorophores and exhibits high selectivity and sensitivity to the c-MYC i-motif in the concentration range of 0.2~1.4 μmol/L, with a detection limit as low as 154 nmol/L (Figure 25b). This probe is capable of rapidly screening selective binding ligands under neutral complex conditions, providing a powerful tool for studying the biological functions of i-motif (Figure 25c).
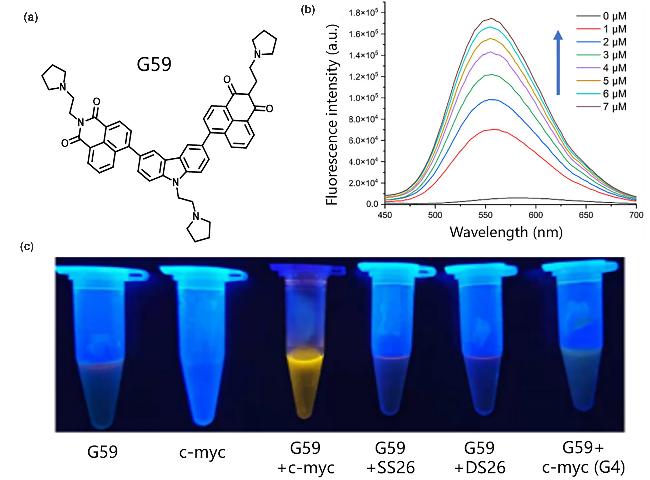
Figure 25 (a) Schematic diagram of the structure of G59; (b) Fluorescence emission spectra of G59 after adding different concentrations of i-motif; (c) Fluorescence changes of G59 binding with i-motif containing c-MYC promoter and other DNAReproduced from [94]. Copy right 2022 MDPI. |
Ruthenium(II) polypyridyl complexes have been extensively studied as potential in vivo fluorescence probes due to their excellent photophysical properties. These compounds exhibit fluorescence-on effect in the presence of DNA. Wallerʼs group[95] investigated the interaction of [Ru- (bqp)2]2+ with the i-motif and other DNA structures (Figure 26a). As shown in Figure 26b, with the addition of DNA, its cis isoform showed a significant fluorescence-on effect up to 50-fold. In addition, the cis isoform indicated the presence of the death-associated protein (DAP) i-motif by fluorescence lifetime measurements, and in the case of mixing with other DNA structures, the lifetime growth of DAP was more significant than that of other types of DNA (Figure 26c). This finding suggests that the potential of steric-diffe- rentiated [Ru(bqp)2]2+ probe in i-motif detection.
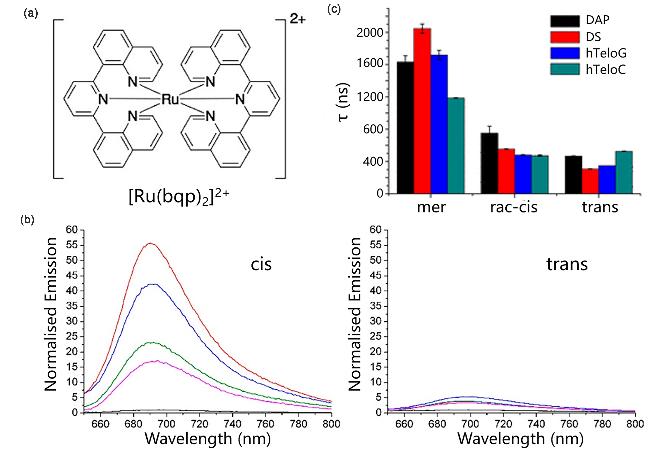
Figure 26 (a) Structurs of [Ru(bqp)2]2+; (b) Normalized emission intensities of cis,trans in the absence of DNA (black), hTeloC (green), DAP (blue), DS (red), and hTeloG (pink); (c) Fluorescence lifetimes of cis, trans and monomers under different conditionsReproduced from [95]. Copy right 2020 American Chemical Society. |
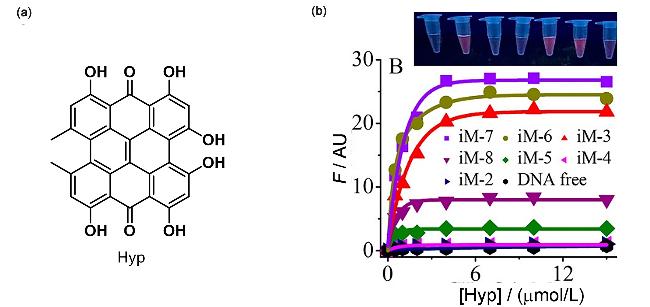
The studies have shown that the length of the human telomere i-motif sequence [5′-(CCCTAA)n-1CCC-5′ (iM-n, n is the number of CCCTAA clusters)] is strongly correlated with the state of aging and health.[96] Shaoʼs group[97] used the natural biotin hypericin (Hyp) as a human telomere i-motif selective probe (Figure 27a). Hyp specifically recognizes C-C base pairs exposed in solution. Binding of Hyp to exposed C-C base pairs is dependent on π-π and cation-π interactions. The enhanced fluorescence of AIE-active Hyp upon binding to the i-motif was able to distinguish i-motif structures of different lengths by fluorescence intensity. Hyp exhibited weak fluorescence upon binding to iM-4, but relative to iM-4, the fluorescence of iM-5, iM-8, iM-3, iM-6, and iM-7 increased 3.8, 9.1, 25.0, 27.6, and 30.1-fold (Figure 27b). This makes Hyp an effective method for detecting the structural polymorphisms of i-motifs.
3.2 Triplex detection
Triplex nucleic acid is a three-dimensional nucleic acid structure formed by the specific binding of triplex-forming oligonucleotide (TFO) to the large groove region of dsDNA through base pairing.[98] Its core feature is that there are two main binding modes between the orientation of the phosphodiester skeleton of TFO and the orientation of dsDNA: when the two skeletons are in the same direction, a parallel triplex is formed, and the bases of TFO are paired with the purine bases of dsDNA (especially A and T) by Hoogsteen hydrogen bond. If the skeleton is reversed, an anti-parallel triplex is formed, but this type of conformation is less stable. At the level of life regulation, Triplex can inhibit or activate transcription processes by binding specific gene promoter regions, thereby regulating key pathways such as apoptosis, DNA repair and cancer progression.[99]
Shaoʼs group[100] synthesized an uncharged NIR probe, NIAD-4, capable of rotational restriction upon binding to Triplex structures, resulting in fluorescence intensifying (Figure 28a). The ability of NIAD-4 to discriminate the Triplex from G4s, i-motifs, and dsDNAs, and its high selectivity is attributed to the topological matching with the Triplex-terminal triplex topology, which enhances its stacking interactions. The same group designed another fluorescence probe called flavonol compound fisetin (FIS), which showed remarkable selectivity and specificity in recognizing Triplex.[101] FIS was able to stabilize the Triplex structure and bind to both ends of Triplex in a 2∶1 molar ratio with a binding constant of approximately (3.6±0.3)×106 L•mol-1. As shown in Figure 28b, FIS triggered the excited-state intramolecular proton transfer (ESIPT) process upon binding to Triplex, generating an intense green fluorescent signal. Whereas, no fluorescence outputted when binding ssDNA, dsDNA, i-motif and G4s, indicating its high selectivity for Triplex structure.
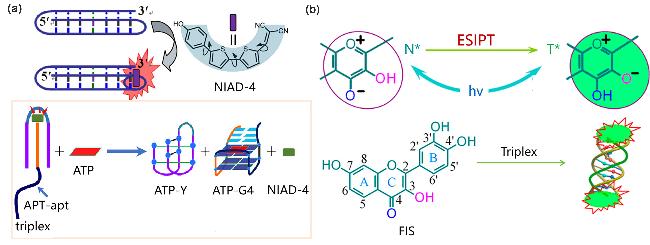
Figure 28 (a) Schematic diagram of Triplex detection mechanism of NIAD-4 and adenosine triphosphate (ATP) sensor based on Triplex; (b) FIS combined with Triplex to trigger ESIPT process to turn on fluorescence.(a) Reproduced from [100]. Copy right 2023 American Chemical Society; (b) Reproduced from [101]. Copy right 2015 American Chemical Society. |
Lingʼs group utilized a fluorescent dye called berberine to identify and detect Triplex structures.[102] Berberine is a plant-derived isoquinoline alkaloid that barely or weakly luminesces in aqueous solution, and when it binds to the Triplex structure, it displays strong fluorescence at 530 nm. In addition, a label-free fluorescence sensing platform was developed for H7N9 viral DNA and thrombin based on the hybridized chain amplification reaction (HCR) and Triplex assembly (Figure 29). When target DNA is present, HCR triggers a hybridization event between two DNA hairpin probes, generating a nicked double helix structure resembling an alternating copolymer. Subsequently, the addition of TFO, which binds to the HCR product to form a stable Triplex structure, resulted in a significant enhancement of berberine fluorescence intensity, with detection limits of 0.14 and 0.32 nmol/L for H7N9 viral DNA and thrombin, respectively.
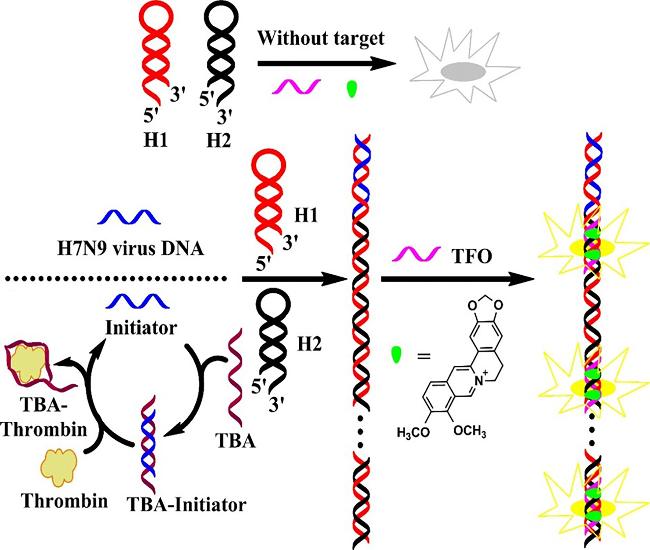
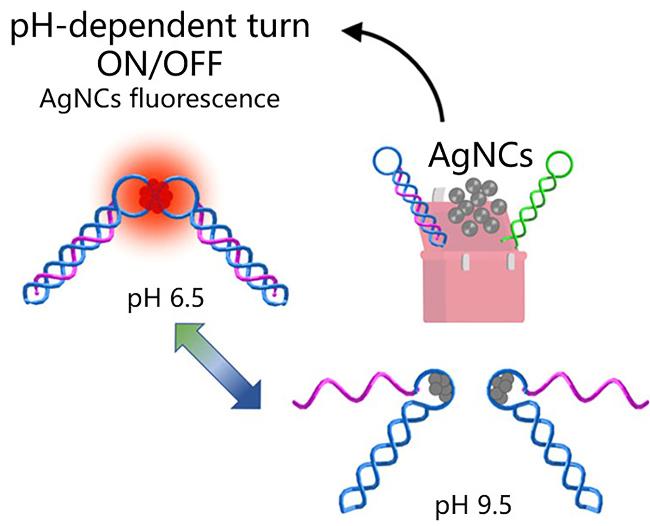
Beside the organic dye, Yangʼs group[103] investigated the binding and fluorescence of inorganic silver nanoclusters (AgNCs) to Triplex using a template of the 10A-10T-7C- 10T Triplex structure containing seven cytosine rings (Figure 30). It was found that the fluorescence of AgNCs embedded in Triplex structures containing heptacytosine rings could be switched on and off with pH varying. Under near-neutral pH conditions (pH 6.5), the Triplex structure was stabilized and AgNCs emitted red fluorescence, while under alkaline conditions (pH 9.5), the Triplex structure dissociated and the fluorescence diminished or disappeared. This dynamic changes in the Triplex structure directly caused changes in the photophysical properties of AgNCs, thus enabling sensitive detection of the Triplex nucleic acid structure.
3.3 Circular nucleic acid (circDNA/RNA) analysis
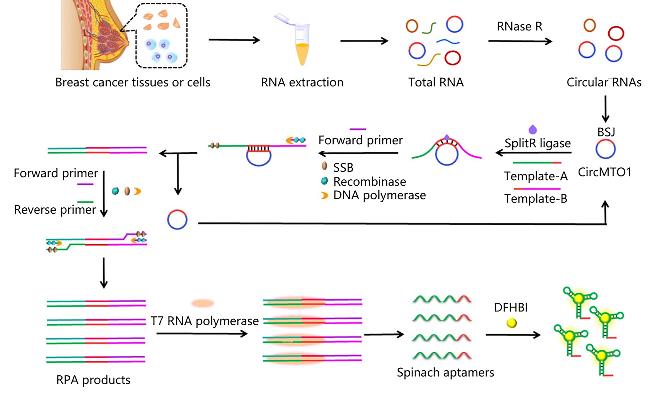
Circular RNAs (circRNAs) are endogenous non-coding RNAs with covalently closed loop structures that are widely found in mammalian cells. Aberrant expression of circRNAs is closely associated with the onset and progression of a variety of diseases (such as metabolic disorders, tumors, and neurological disorders). Zhangʼs group developed a biosensor based on proximal ligation-activated recombinase polymerase amplification (RPA) and transcriptional amplification technologies for the detection of circRNAs (Figure 31).[104] The method used circMTO1 as a template and induced a proximal ligation reaction to activate RPA, generating dsDNA containing the T7 promoter, which is subsequently transcriptionally amplified by the T7 RNA polymerase to produce a large amount of Spinach RNA aptamer. This aptamer specifically bound to DFHBI (3,5-difluoro-4-hydroxybenzylidene imidazolidinone) dye and produced a significant fluorescence signal with a limit of detection of 2.54×10-18 L•mol-1. Importantly, the expression of circ- MTO1 was found to be lower in tissues of breast cancer patients than that from healthy individuals, indicating that the abnormal expression of circMTO1 is closely related to breast cancer.
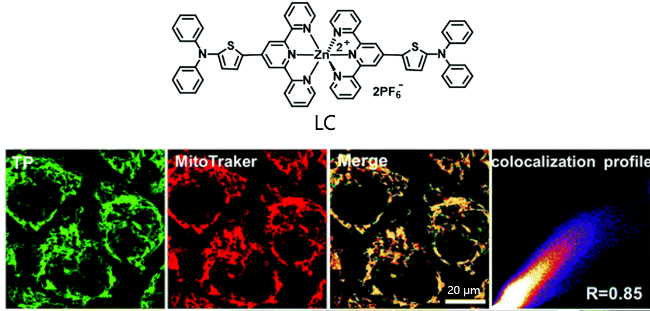
Mitochondrial DNA (mtDNA) is a typical closed-loop double-stranded structure of nucleic acids, which is an important genetic material encoding 37 genes and involved in many biological functions. Unlike nuclear DNA, mtDNA lacks histone protection and endonuclease repair systems, and the RNA primer-mediated strand elongation mechanism during replication significantly increases its risk of exposure to highly oxidizing environments, resulting in a mutation rate up to 10-100-fold that of nuclear DNA.[105] This unique molecular stability defect makes it an ideal biomarker for monitoring oxidative damage in cells. Dete-ction of mtDNA will be useful in assessing the health of cells and organisms as well as in the diagnosis of mtDNA- associated diseases. Tianʼs group[106] thus synthesized a thiophen-based tripyridine Zn(Ⅱ) complex LC, whose remarkable planarity of LC enables its insertion into the double helical structure of mtDNA. Its binding to mtDNA was attributed to the π-π interactions with the large π- bonding system in the bases and hydrophobic interactions, with a binding constant of 7.04×105 L•mol-1. Particularly, LC exhibited efficient intracellular uptake and overlapped with the fluorescence of Mito Tracker Deep Red, indicating that LC was able to localize in mitochondria, enabling high- resolution imaging of mtDNA in live cells (Figure 32).
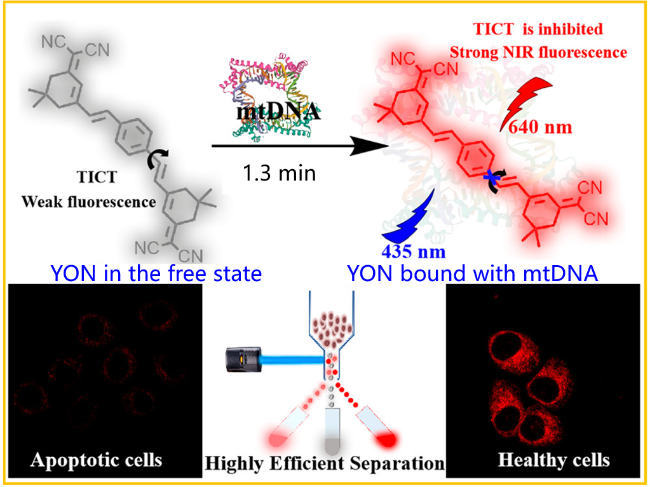
Zhangʼs group[107] synthesized a near-infrared twisted intramolecular charge transfer (TICT)-based fluorescence probe YON for mtDNA detection (Figure 33). With a strong electronic push-pull and TICT properties, YON is able to bind mtDNA tightly by embedding small grooves with a binding constant of 8.5×105 L•mol-1, leading to an ultra-sensitive fluorescence signal response at 640 nm with a detection limit of 71 ng/mL. The Stokes shift of over 200 nm and the near-infrared fluorescence emission wavelength enable YON to specifically and highly sensitively monitor the small changes of mtDNA in complex biological environments and assess cellular health in real time manner.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In this paper, probes targeting i-motif recognize the target structure through π-π stacking and protonation response mechanism, achieving dynamic monitoring of pH-sensitive conformations and providing specific tools for tumor acidic microenvironment studies and telomere function regulation;while triplex probes not only achieve highly selective imaging of Triplex, but also have been expanded to the fields of virus detection and ATP sensing; and cyclic nucleic acid probes have shown unique advantages in circRNA hypersensitivity detection and mtDNA oxidative damage assessment. Currently, these probes have broken through the limitations of in vitro detection and have been gradually applied to live cell imaging and disease marker screening, but still need to solve the technical bottlenecks such as insufficient tissue penetration depth and simultaneous identification of multiple targets. In the future, by integrating NIR-III optical window, DNA nanotechnology and single-molecule imaging, atypical nucleic acid probes are expected to achieve breakthroughs in the fields of tumor heterogeneity analysis, aging mechanism investigation and mitochondrial disease diagnosis and treatment. Particularly, the association of conformational dynamics monitoring with epigenetic regulation, metabolic reprogramming and other mechanisms may reveal new disease targets, promote the developments of precise medicine based on nucleic acid structure regulation, and ultimately realize the whole chain of innovation from molecular mechanism to clinical translation.
4 Summary and prospects
Fluorescence probes for detection of atypical nucleic acid structures had made remarkable progress in recent years. The target is to reveal the expression level, dynamic conformation and biological functions of various atypical nucleic acid structures and to promote their applications in disease diagnosis and therapy. Currently, multi-dimensional strategies have been formed for probe design, and various types of fluorescence molecules can realize specific recognition of atypical nucleic acid structures through the introduction of motifs that can generate π-π stacking, electrostatic attraction or hydrogen bonding, etc. (Table 1). Fluorescence turn-on probes have significant advantages in imaging atypical nucleic acid structures, mainly including high signal-to-noise ratio and low background signal, which enable them to sensitively detect low-abundance targets and are suitable for real-time dynamic monitoring of biotargets in living cells and living bodies. In addition, through the target-induced fluorescence turn-on and/or enhancement mechanism, these probes can effectively reflect the dynamic conformational transitions of nucleic acid structures (such as G-quadruplex, i-motif, etc.), which provides an important tool for studying their biological functions. Combined with ratiometric fluorescence design, some probes can also improve the accuracy of quantitative analysis. The aggregation-induced emission luminogens (AIEgens) further improves photostability and resistibility to photobleaching, while the modular probe design combined with nucleic acid aptamer technology enables highly specific detection at the single-molecule level.
Table 1 Comparison of different fluorescent probes for the detection of atypical nucleic acids |
| Probe | Types of nucleic acid | Key features | Ref. |
|---|---|---|---|
| CSTS | G4s | Disaggregation-Induced Emission (DIE), V-shaped rigid plane π bracket combined with the parallel G4s through the | [42] |
| QCy(BnBT)3 | G4s | High specificity recognition of G4s and 500 times fluorescence enhancement | [43] |
| SCY-5 | G4s | J-Aggregates dissociate into monomers, high specificity recognition of parallel G4s | [44] |
| SN-Cy5-S | G4s | Disaggregation-Induced Emission (DIE), Y-shaped plane is combined with the top plane at the 5' end of G4s | [45] |
| TO | G4s and Triplex | Distinguish between Triplex and G4s, strong fluorescence enhancement | [47] |
| TOVJ | G4s | High specificity recognition of antiparallel G4s, near-infrared emission | [48] |
| TOR-G4 | G4s | Two-photon excitation, fluorescence lifetime has increased significantly | [49] |
| ThT | G4s | High cell membrane permeability, specific recognition ability for G4s | [50] |
| IMT | G4s | Stack on the 3'-G4 end plane, low background and high signal-to-noise ratio emissions | [51] |
| ThT-NE | G4s | Intramolecular rotation is restricted and fluorescence is activated, high selectivity and sensitivity | [52] |
| ThT-NA | G4s | Red light emission, large Stokes shift, high fluorescence turn-on ratio and high selectivity for G4s | [53] |
| NIRG-2 | G4s | Near-infrared Region II emission, hydrogen bonds and π-π stacking combine with G4s | [54] |
| CQ4 | G4s | Disaggregation-Induced Emission (DIE), specifically bind parallel G4s | [55] |
| P0 | G4s | The dimer G4/P0 system carried out highly selective detection of K+ | [56] |
| SiR-PyPDS | G4s | The G4s ligand Pyridostatin (PyPDS) forms hydrogen bonds and hydrophobic interactions to specifically bind to G4s | [57] |
| DAOTA-M2 | G4s | G4s were identified by fluorescence lifetime imaging microscopy (FLIM) technology | [58] |
| SG4 | G4s | Antibodies against the structure of human c-MYC G4, bind to green fluorescent protein (GFP) | [59] |
| CV2 | G4s | Peptide sequences specifically recognizing G4s (L-ARG-L-Gli-glutaric acid), Imaging of mitochondrial G4s | [60] |
| COL | G4s | Induce parallel G4s to aggregate from the nucleic acid mixture | [61] |
| TTAPE | G4s | Aggregation-induced emission detection (AIE), real-time monitoring of the folding process of G-rich DNA strands to form G4s | [84] |
| PZ-1 | G4s | Aggregation-induced emission detection (AIE), G4s is combined through electrostatic interaction and π-π stacking | [85] |
| TPE-mTO | G4s | Electron-deficient cation conjugated systems and crescent-shaped scaffolds, specifically locate G4s in mitochondria | [86] |
| TPA-mTO | G4s | Near-infrared emission, ideal photostability, and high fluorescence contras | [87] |
| HMPQ | G4s | AIEgens of biological origin, the π-π interaction binds to G4s, high selectivity and high sensitivity | [88] |
| 5i and 5c | i-motif | Combining different types of I-motifs, the fluorescence intensity is enhanced and the fluorescence lifetime is increased | [93] |
| G59 | i-motif | Specifically recognize and visually detect the i-motif structure of the c-MYC gene promoter | [94] |
| [Ru(bqp)2]2+ | i-motif | The cis isomer combines with the DAP i-motif, resulting in fluorescence activation and increased fluorescence lifetime | [95] |
| Hyp | i-motif | The i-motif structures of different lengths were distinguished by fluorescence intensity | [97] |
| NIAD-4 | Triplex | Uncharged near-infrared molecular rotor probe, topological match with the Triplex terminal triad | [100] |
| FIS | Triplex | The quantity ratio of 2∶1 is combined with Triplex, the ESIPT process is triggered and the green fluorescence lights up | [101] |
| Berberine | Triplex | Isoquinoline alkaloids, combined with the Triplex structure, it will show strong fluorescence at 530 nm | [102] |
| AgNCs | Triplex | The dynamic changes of photophysical properties with the Triplex structure | [103] |
| DFHBI | circRNA | Using circMTO1 as the template, the aptamers generated by RPA and transcriptional amplification techniques were combined with DFHBI | [104] |
| LC | circDNA | Efficient intracellular uptake capacity, high-resolution visual imaging detection of mitochondrial DNA (mtDNA) | [106] |
| YON | circDNA | Near-infrared twisted intramolecular charge transfer (TICT) fluorescent probe, high sensitivity and large Stokes displacement | [107] |
Despite the remarkable progresses achieved in the life sciences, multiple challenges still exist. For instance, the selectivity, metabolic stability, tissue penetration depth, phototoxicity, and multi-target simultaneous detection ability of probes still need to be improved. The “turn-on” state of the probe is susceptible to microenvironmental factors (such as pH, viscosity, polarity, etc.), which can lead to false positives. In addition, probe design requires a precise balance between on/off ratio and binding affinity, and over-optimization may impair dynamic monitoring. In complex biological systems, non-specific binding and cross-reactivity may reduce detection specificity, while strong binding may even interfere with the natural conformational equilibrium of the target structure, leading to observation errors. Currently, most turn-on probes are still limited to the detection of specific sequence backgrounds and lack the panoramic imaging capability of global atypical nucleic acid structures, and their metabolic stability, tissue enrichment efficiency, and potential phototoxicity in in vivo applications still need to be further optimized. Together, these factors constrain the widespread use of this technology in in vivo research and clinical translation.
To address these challenges, future research needs to make breakthroughs in multiple dimensions. At the technical level, the development of multimodal probes, combined with fluorescence, Raman and magnetic resonance imaging, can realize the simultaneous monitoring of nucleic acid structure and microenvironmental parameters, and improve the accurate recognition ability of probes. Meanwhile, artificial intelligence-assisted molecular design will be used to optimize the structure-property relationship of the probes and enhance the selectivity for specific topological conformations (such as antiparallel G4s). In addition, promoting the deep integration of NIR-II and AIEgens technology, and designing probes with deep tissue penetration, low background noise, and real-time dynamic response will provide a powerful tool for the visualization of metastatic tumors or neurodegenerative lesions in vivo. At the application level, targeted delivery systems (such as tumor-derived exosomes) can enhance the efficiency of probe focal enrichment, while the nucleic acid conformation-gene expression association mapping established by combining single-cell sequencing and spatial genomics technologies will lay the molecular foundation for precision medicine. Particularly, it is crucial to strengthen the clinical translational research of probes. Through the development of targeted delivery systems (such as exosomal carriers), the enrichment efficiency of probes in the lesion site can be improved, which will help realize precise medicine. Meanwhile, combining single-cell sequencing and spatial genomics technologies to establish spatial correlation maps of nucleic acid conformation and gene expression will provide a new molecular basis for disease diagnosis and treatment. In addition, exploring the cross-regulatory mechanism of non-classical nucleic acid structure with epigenetic modification and metabolic reprogramming is expected to reveal new targets for disease treatment and promote the development of novel drug-like molecules and smart fluorescence probes.
In summary, the research of atypical nucleic acid fluorescence probes is in the stage of rapid development, and is gradually breaking through the boundaries of diagnosis and treatment through interdisciplinary collaboration and technological innovation. In the future, these probes are not only expected to reveal the molecular mechanism of life activities, but also may lead to the discovery of new targets for drug therapy and the development of new diagnostic and therapeutic strategies based on the dynamic conformation of nucleic acids, which will become the next generation of molecular tools to analyze the mysteries of life and revolutionize the diagnosis and treatment of diseases. With the continuous progress of technology, fluorescence probe technology will play an increasingly important role in the field of life sciences.
(Lu, Y.)